Навигация
Влияние кинетина на физиологические процессы
58703
знака
17
таблиц
11
изображений
4.2 Влияние кинетина на физиологические процессы
Одним из проявлений действия фитогормонов является усиление роста стебля в высоту. Поэтому прежде всего нами проводились исследования по влиянию кинетина на рост растений пшеницы.
Данные по этому вопросу представлены в таблицах 1 и 2.
Таблица 1 Влияние кинетина на длину побеговой системы 10-ти проростков пшеницы ( см).
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 7 | 15,4±0,15 | 14,64±0,3 | 95 | 16,1±0,39 | 15,83±0,17 | 98 |
| 9 | 15,9±0,66 | 15,7±0,66 | 99 | 17,4±0,21 | 16,4±0,39 | 94 |
| 11 | 18,1±0,12 | 17,9±0,09 | 99 | 18,1±0,75 | 17,7±0,18 | 98 |
| 16 | 18,5±0,15 | 18,2±0,42 | 98 | 20,6±0,36 | 18,3±0,36 | 89 |
Таблица 2 Влияние кинетина на длину корневой системы 10-ти проростков пшеницы ( см )
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 7 | 14,8±0,24 | 14,0±0,33 | 95 | 14,96±0,16 | 14,9±0,06 | 100 |
| 9 | 15,6±0,6 | 15,1±0,12 | 97 | 15, 5±0,17 | 15,1±0,07 | 97 |
| 11 | 17,5±0,1 | 15,51±0,18 | 89 | 16,08±0,17 | 15,32±0,17 | 95 |
| 16 | 17,85±0,24 | 16,1±0,13 | 90 | 16,65±0,13 | 15,9±0,21 | 95 |
Как видно из таблиц 1 и 2 кинетин практически не повлиял на длину корневой и побеговой системы.
Учитывая то, что калий не повлиял на длину побеговой системы, но вызвал увеличение ее массы, мы провели исследования по влиянию кинетина на накопление массы корневой и побеговой систем в зависимости от уровня минерального питания. Данные расположены в таблицах 3 и 4.
Таблица 3 Влияние кинетина на массу побеговой системы 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мг).
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 7 | 0,830±0,022 | 0,843±0,024 | 102 | 0,886±0,051 | 0,907±0,098 | 102 |
| 9 | 0,904±0,057 | 0,922±0,101 | 102 | 1,056±0,106 | 1,234±0,092 | 117 |
| 11 | 1,094±0,025 | 1,259±0,083 | 115 | 1,409±0,095 | 1,540±0,078 | 109 |
Таблица 4 Влияние кинетина на массу корневой системы 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мг)
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 7 | 760±3 | 681±18 | 90 | 740±17 | 742±8 | 100 |
| 9 | 770±18 | 741±76 | 96 | 796±8 | 770±17 | 97 |
| 11 | 830±46 | 996±10 | 120 | 822±58 | 825±68 | 100 |
| 16 | 983±11 | 1030±28 | 105 | 1014±32 | 1051±36 | 104 |
Анализируя таблицы 2 и 3 можно сделать вывод, что кинетин практически не повлиял на массу побеговой и корневой системы. Хотя можно отметить незначительный рост массы побеговой системы у 11-ти дневных проростков.
Как и в опытах с калием, влияние кинетина изучалось и на интенсивность дыхания и фотосинтеза. Данные, полученные в результате опыта приведены в таблицах 5 и 6.
Таблица 5 Влияние кинетина на интенсивность дыхания 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мкл O2 / гч)
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 4 | 450±68 | 510±74 | 113 | 530±125 | 605±130 | 114 |
| 6 | 678±81 | 755±44 | 111 | 948±15 | 1040±10 | 110 |
| 8 | 949±165 | 903±233 | 95 | 999±216 | 1075±227 | 108 |
| 10 | 1500±66 | 1648±44 | 110 | 1720±53 | 1833±26 | 107 |
Таблица 6 Влияние кинетина на интенсивность фотосинтеза 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мкл O2 / гч).
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 4 | 620±83 | 708±4 | 114 | 685±15 | 739±25 | 108 |
| 6 | 698±76 | 720±138 | 103 | 734±79 | 821±286 | 112 |
| 8 | 948±166 | 1180±107 | 125 | 999±216 | 1775±133 | 178 |
| 10 | 1500±144 | 1538±126 | 102 | 1720±120 | 2220±159 | 129 |
И в таблицах 7 и 8 приведены результаты опытов по влиянию кинетина на содержание в проростках сахаров и хлорофилла.
Таблица 7 Влияние кинетина на содержание хлорофилла 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мг/ч).
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 4 | 0,113±0,11 | 0,207±0,11 | 183 | 0,268±0,01 | 0,481±0,042 | 179 |
| 6 | 0,478±0,05 | 0,577±0,04 | 121 | 0,547±0,04 | 0,620±0,04 | 113 |
| 8 | 0,325±0,02 | 0,434±0,02 | 134 | 0,419±0,003 | 0,476±0,005 | 114 |
| 10 | 0,389±0,01 | 0,487±0,013 | 125 | 0,429±0,01 | 0,494±0,01 | 115 |
Таблица 8 Влияние кинетина на содержание сахаров 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мг/г сырого веса)
| Возраст, дн. | NP | NP + КН | % | NPK | NPK + КН | % |
| 4 | 5,614±1,5 | 4,533±1,23 | 81 | 3,765±0,88 | 2,211±0,4 | 59 |
| 6 | 0,374±0,24 | 0,430±0,32 | 115 | 0,845±0,03 | 0,889±0,01 | 105 |
| 8 | 1,167±0,2 | 1,487±0,2 | 127 | 0,959±0,01 | 1,148±0,03 | 120 |
| 10 | 0,770±0,1 | 0,886±0,1 | 115 | 0,983±0,04 | 1,252±0,02 | 127 |
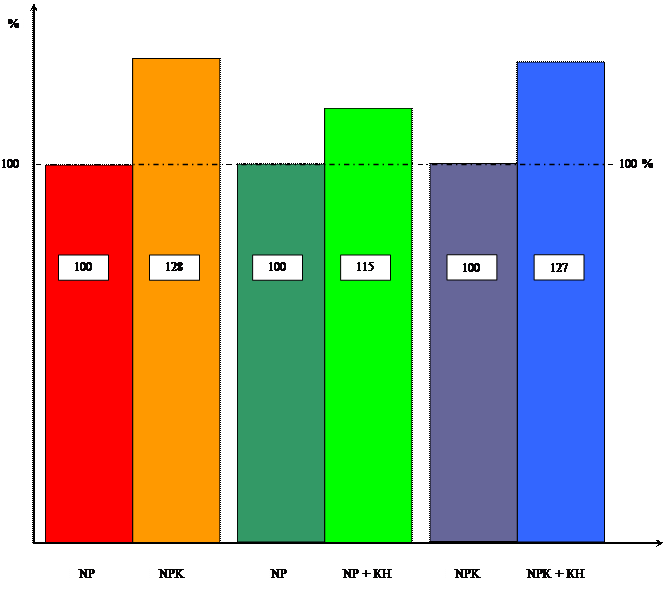
Аналогичная картина наблюдается и по уровню содержания сахаров и хлорофилла, которое под влиянием кинетина тоже увеличивается.
Содержание сахаров в отсутствии калия увеличивается на 16%, а на ППС на 17%. Хотя у проростков в возрасте 4-х дней содержание сахаров уменьшается, т.к. при переходе от гетеротрофного питания к автотрофному усиливается дыхание проростков. Это хорошо видно в таблице 5. В возрасте 6, 8,10 дней содержание сахаров под влиянием кинетина начинает увеличиваться, т.к. увеличивается интенсивность фотосинтеза.
Содержание хлорофилла в отсутствии калия увеличилось на 41%, а на ППС на 30%.
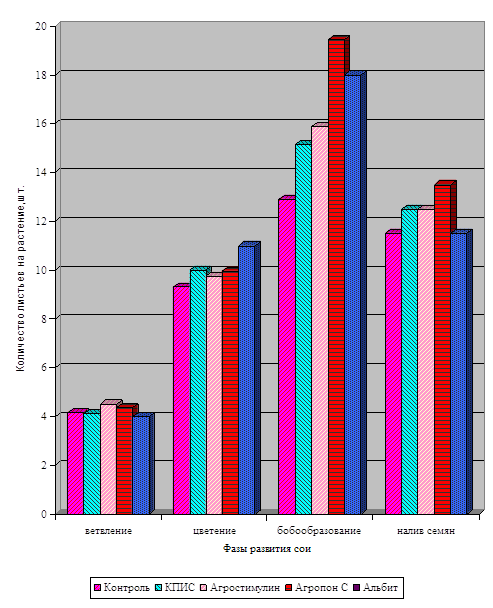
Диаграмма влияния цитокинина и калия
на длину побеговой системы
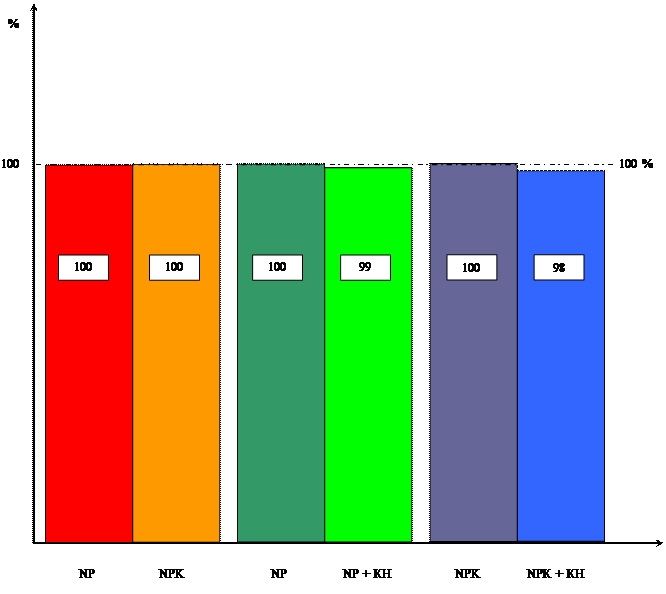
Диаграмма влияния цитокинина и калия
на массу побеговой системы
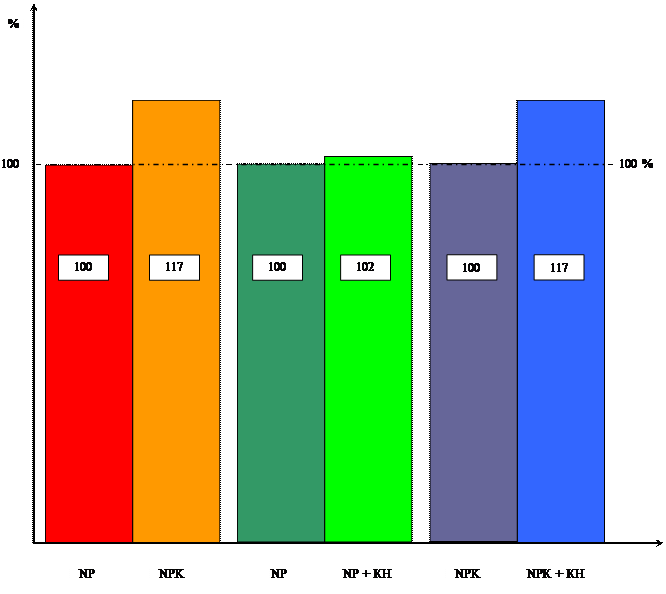
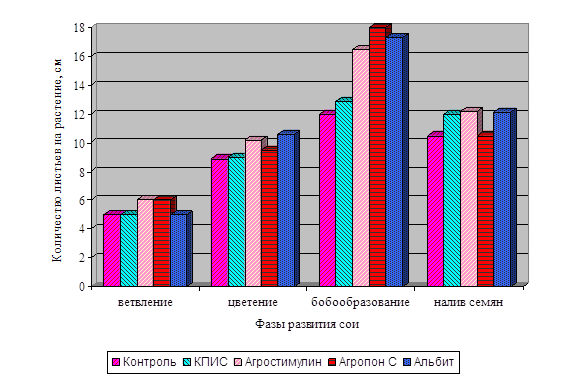
Диаграмма влияния цитокинина и калия
на длину корневой системы

Диаграмма влияния цитокинина и калия
на массу корневой системы
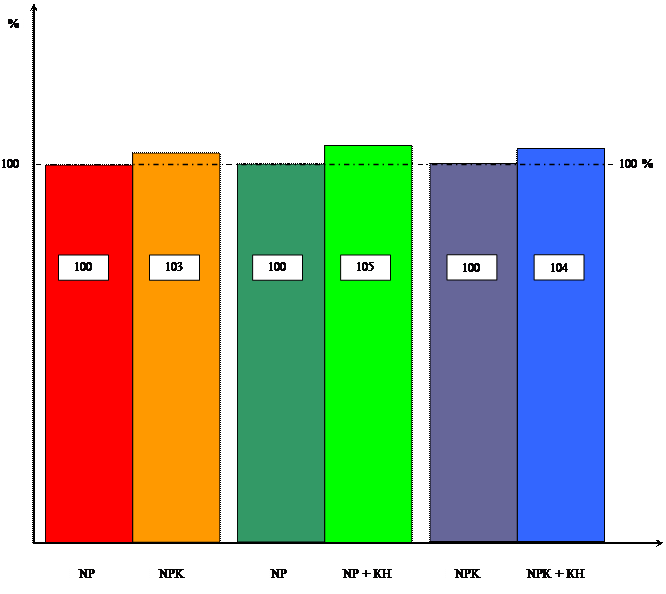
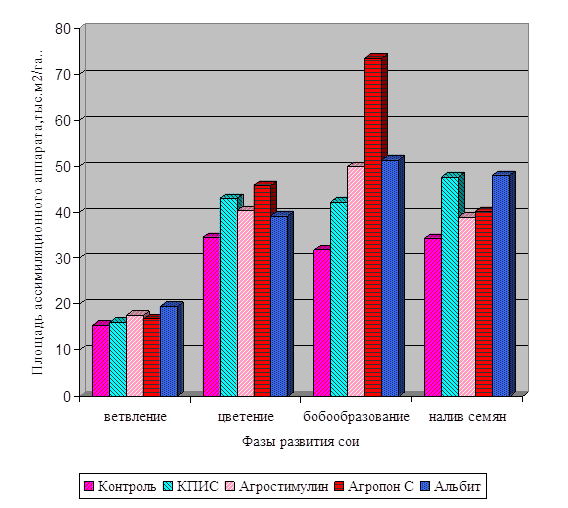
Диаграмма влияния цитокинина и калия
на интенсивность дыхания
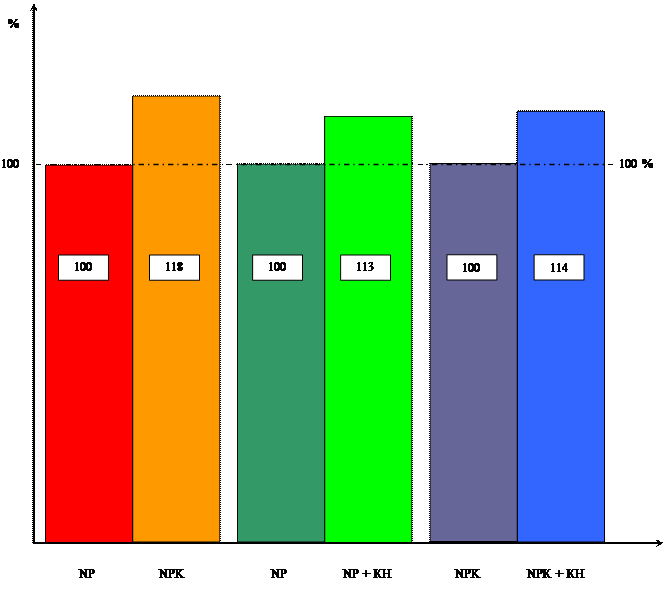
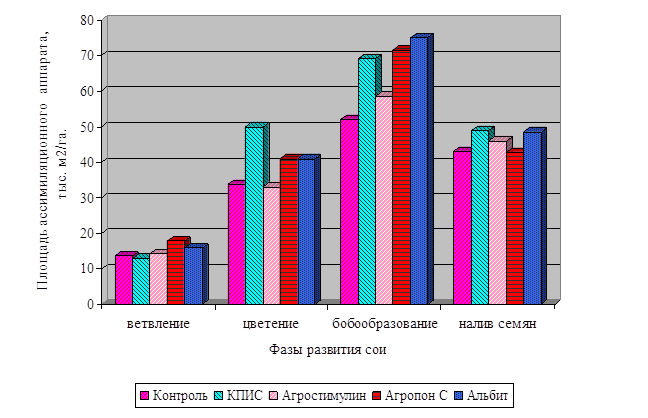
Диаграмма влияния цитокинина и калия
на интенсивность фотосинтеза

Диаграмма влияния цитокинина и калия
на содержание хлорофилла
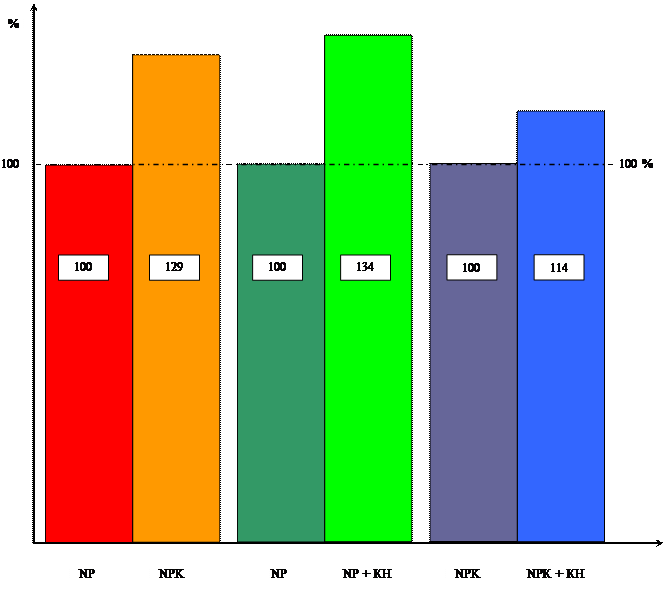
Диаграмма влияния цитокинина и калия
на содержание сахаров
4.3 Выводы
1. Рост растений пшеницы на ППС максимальный. Исключение калия из питательной смеси практически не приводит к изменению роста растений и снижению массы, т.к эти изменения очень не значительны. Влияние калия в ПС сказывается только на увеличении массы надземной части проростков.
2. Под влиянием кинетина увеличение накопления массы проростков практически не наблюдается. В то же время присутствие кинетина значительно влияет на увеличение интенсивности дыхания и фотосинтеза, содержания сахаров и хлорофилла.
3. Влияние кинетина оказывается наиболее эффективным при недостатке калия, что вероятно связано с тем, что кинетин компенсирует недостаток калия в питательной среде. Отсюда можно сделать предположение, что кинетин и калий возможно влияют на одни стороны изучаемых процессов.
5. СПИСОК ЛИТЕРАТУРЫ
1. Артамонов В.И. “Учебные записки МОПИ им.Крупской”, 1967.
2. Артамонов В.И. “Физиология растений”, 1966.
3. Ващенко И.М. “Основы сельского хозяйства”, 1987.
4. Ветшина В.В. “Влияние стимуляторов роста и микроэлементов на урожай карликовых бобов” // Бюл. Гл. ботан. сада, 1963.
5. Верзилов В.Ф. “Регуляторы роста и их применение в растениеводстве”, М., Наука, 1971.
6. Гамбург К.З. “Регуляторы роста и рост растений”. // М., Наука,1964.
7. Гамбург К.З. “Фитогормоны и клетки”. // М., Наука, 1970.
8 . Головко Т.К. “Дыхание растений. Физиологические аспекты”.
9 . Дерфлинг К. “Гормоны растений”, 1985.
10. Доклады РАСХН, 1997 №2 с 17-18.
11. Доклады АН РАН, 1997 т.353 №2 с 261-263.
12. Жарикова А.М. “Питание яровой пшеницы калием из удобрений и почвы”. Весн. АН БССР, 1970.
13. Заманова А.П. “Эффективность применения орг. и мин. удобрений совместно с цитокининами на рост, развитие и урожайность виноградной лозы”. Автореф. диссерт. ... канд. биол. наук.
14. Зединг Г. “ Ростовые вещества растений”, М., изд-во Иностранная литература, 1955.
15. Кулаева О.Н. “Цитокинины, их структура и функции”. М., Наука, 1973.
16. Кулаева О.Н. “Гормональная регуляция физиологических процессов у растений на уровне синтеза РНК и бнлка”. М., 1982.
17. Морозов А.С. “Физиологические основы питания сельскохозяйственных растений”. // Сборник статей - М., 1983.
18. Маховская М.А. “Влияние физиологически активных веществ на фотосинтез”. // Автореф. канд. дис. ... Киев, 1965.
19. Мамедов З.И. “Питание и урожай с/х растений”. АН Азербайджан, 1991 год.
20. Овсянникова Е.Н. “Особенности гормональной регуляции ростовых поцессов у растений огурца”, В сб.: Физиологические основы ростовых процессов, М., МОПИ, 1986.
21. Петербургский А.В. “Агрохимия и физиология питания растения”.
изд. 2-е, перераб. - М., Россельхозиздат, 1981.
22. Похлебаев С.М. “Изменение функциональной активности хлоропластов ячменя и пшеницыпод действием цитокинина”. Доклады ВАСХНИЛ, 1981.
23. Полевой В.В. “Фитогормоны”. Л., ЛГУ, 1982.
24. Полевой В.В. “Физиология растений”. М., Высшая школа, 1989.
25. // Прикладная биохимия и микробиология, 1993, т29 вып. 5
с 644-674.
26. // Прикладная биохимия и микробиология, 1994, т30 вып. 2 с 284-291.
27. Пушкина Г.П. “Влияние гибберелина икинетина на процесс синтеза и разрушения хлорофилла в проростках кукурузы”, М., МОПИ, 1973.
28. Ростунов А.А. “Влияние фитогормонов на поступление ионов в зависимости от уровня минерального питания”. // Межвузовский сборник научных трудов, МОПИ, 1988.
29. Ростунов А.А. “Влияние азотного питания и фитогормонов на физиологические процессы и рост двух сортов озимой пшеницы разной продуктивности”. М., 1990.
30. Соколов А.В. “Агрохимия калия”. М., АН СССР, 1950.
31. Ситникова О.А., Гриненко О.А., Силкова Л.Г. “ Регуляторы роста и их действие на растения”. М., МОПИ, 1967.
32. Сивцова А.М. “Рольфитогормонов в реакции растений на уровень минерального питания”. // Автореф. дис. ... канд. биол. наук, / ТСХА - М., 1993.
33. // Физиология растений, 1993 т40 №6 с 893-897.
34. // Физиология и биохимия культурных растений,1995 т27 №1-2 с 31-35.
35. // Физиология и биохимия культурных растений, 1997 т29 №3 с 220-225.
36. // Физиология растений, 1994 т41 №3 с 448-459.
37.// Физиология растений, 1982 т29 вып.3 с 502-507.
38. Физиология и биохимия культурных растений, 1999 т31 №1 с 53-60.
39. Хвойка Людовит “Цитокинины и ауксины в регуляции роста растений”.
40. Хоменко А.Д. “Корневое минеральное питание и продуктивность растений”. Киев, Наукова думка, 1976.
41. Чайлахян М.Х., Некрасова Т.В. “Влияние веществ высокой физиологической активности на преодоление полярности у черенков лимона”. М., ДАН СССР, 1958.
42. Чуйкова Л.В. “Особенности физиологического действия регуляторов роста при опрыскивании полевых культур в целях повышения их продуктивности”. Автореф. дис. ... канд. биол. наук. Ворлнеж, 1964.
43. Чуйкова Л.В. “Влияние регуляторов роста на физиологические процессы и урожай сахарной свеклы”. // Регуляторы роста растений, Воронеж, 1964.
44. Чуйкова Л.В. “Влияние регуляторов роста на физиолого-биохимические процессы и продуктивность кукурузы”, В сб.: Регуляторы роста растений, Воронеж, 1964.
45. Эрдели Г.С. “Влияние регуляторов роста на продуктивность и некоторые физиологические процессы подсолнечника”. В сб. Регуляторы роста растений, Воронеж, ВГУ, 1964.
46. Якушкина Н.И. “Повышение эффективности минеральных удобрений при помощи регуляторов роста”. // Физиологическое обоснование системы питания растений, М., НаукаБ 1964.
47. Якушкина Н.И. “Фитогормоны и их действие на растение”. М., 1982 год.
48. Якушкина Н.И. “Физиология растений”, М., Просвящение, 1980.
49. Якушкина Н.И., Похлебаев С.М. “Особенности фосфорилирования хлоропластов, выделенных и обработанных фитогормонами листьев ячменя и пшеницы”, Физиология растений т29, вып. 3, 1982.
50. Якушкина Н.И., Пушкина Г.П. “Изменение интенсивности фосфорилирования в проростках кукурузы под действием гибберелина и кинетина”, Физиология растений т22, вып. 6, 1975.
Похожие работы
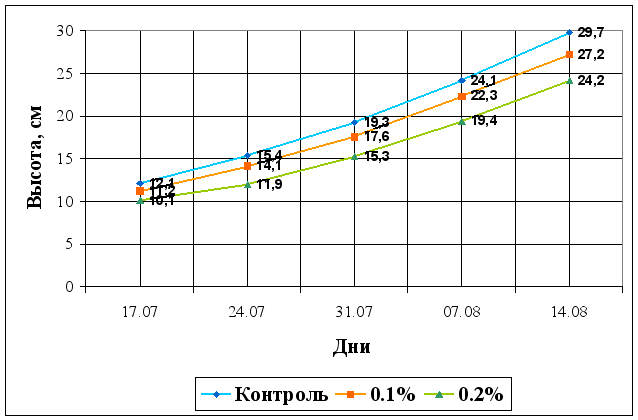
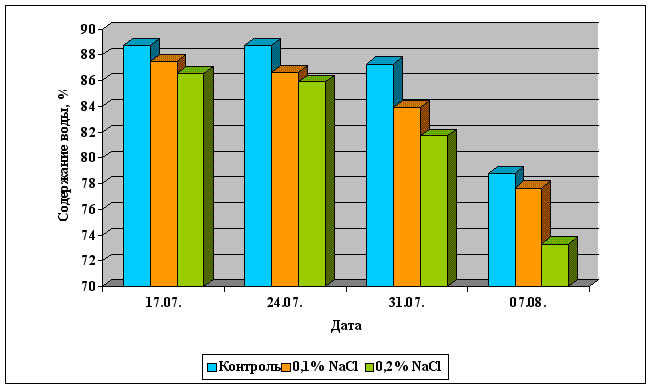
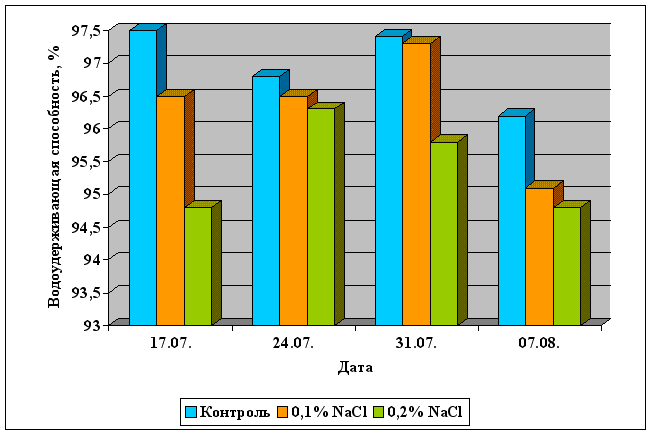
... Областного Университета в период с июня по сентябрь 2003 года. Объектом изучения являлись растения кукурузы сорта Россо. Изучение влияния 6-БАП на рост и биометрические показатели растений кукурузы при разном уровне засоления проводилось в условиях вегетационного опыта (почвенные культуры). Семена кукурузы проращивались при температуре 20оС в термостате в течение 3 дней, а затем высаживались в ...
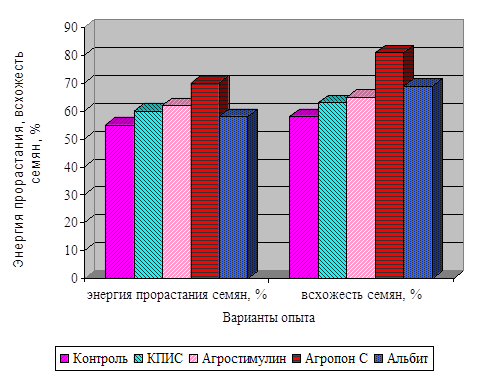
... и болезней. Но эти методы в основном негативно сказываются на качестве продукции и на окружающей среде. В наших исследованиях по изучению влияния регуляторов роста растений на физиолого-биохимические показатели и урожайность сои мы применяли только экологически безопасные препараты. Это продукты жизнедеятельности микроорганизмов и идентичные по своему происхождению природным соединениям, ...
... пшеницы сорта эритроспермум 59 Проведенные исследования (таблица 10) свидетельствуют о том, что наименьший урожай яровой пшеницы Эритроспермум 59 наблюдается на безгербицидном варианте, без Гуми-М (1,88 т/га). Таблица 10 - Влияние обработок посевов гербицидом Пума супер 100 и адаптогенным препаратом на урожайность яровой пшеницы (2001-2003 гг.) Вариант (норма расхода гербицида, л/га) ...
... гербицид нарушает полярность, вызывает утолщение побегов, опадение листьев, морфозы, в результате жизнедеятельность растений дезорганизуется и наступает их гибель. Ретарданты. Это синтетические регуляторы, тормозящие биосинтез гиббереллинов, подавляющие рост стебля и вегетативных побегов, придающие растениям устойчивость к полеганию. В то же время они способствуют росту корней, листьев, повышают ...
0 комментариев