Навигация
Содержание аскорбиновой, дегидроаскорбиновой и дикетогулоновой кислот в эритроцитах здоровых детей и страдающих инсулинзависимым сахарным диабетом
41601
знак
1
таблица
7
изображений
МИНИСТЕРСТВО ОБЩЕГО И ПРОФЕССИОНАЛЬНОГО
ОБРАЗОВАНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
КРАСНОЯРСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Биологический факультет
Кафедра биохимии и физиологии человека и животных
С.А. Костогорова
студентка 4 курса
СОДЕРЖАНИЕ АСКОРБИНОВОЙ, ДЕГИДРОАСКОРБИНОВОЙ И ДИКЕТОГУЛОНОВОЙ КИСЛОТ В ЭРИТРОЦИТАХ ЗДОРОВЫХ ДЕТЕЙ И СТРАДАЮЩИХ ИНСУЛИНЗАВИСИМЫМ САХАРНЫМ ДИАБЕТОМ
(курсовая работа)
Научный руководитель:
к.б.н., доц. Титова Н.М.
![]()
Красноярск, 1999
оглавление
Введение..................................................................................................... 2
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ................................................................. 3
1.1.биохимические процессы при созревании и старении эритроцитов.... 3
1.1.1. Характеристика эритроцитов.......................................................... 3
1.1.2. Энергетический обмен в эритроцитах............................................ 5
1.1.3. Антиоксидантная система эритроцитов.......................................... 6
1.2. Аскорбат как компонент АОС эритроцитов........................................ 8
1.2.1. Строение и физико-химические свойства аскорбата..................... 8
1.3. Сахарный диабет как один из распространёенных патологических процессов 9
ГЛАВА 2. МАТЕРИАЛЫ И МЕТОДЫ....................................................... 11
2.1. Подготовка эритроцитов..................................................................... 11
2.2. Метод раздельного определения аскорбиновой, дегидроаскорбиновой и дикетогулоновой кислот в эритроцитах.................................................... 11
2.3. СТАТИСТИЧЕСКАЯ ОБРАБОТКА РЕЗУЛЬТАТОВ......................... 13
Глава 3. РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ....................................... 14
ВЫВОДЫ...................................................................................................... 15
литература.............................................................................................. 16
SUMMARY.................................................................................................... 19
Приложение................................................................................................... 20
Введение
Зрелые эритроциты млекопитающих – это высокоспециализированные безъядерные клетки. Основной функцией эритроцитов является транспорт кислорода от клетки к тканям и углекислоты в обратном направлении. Высокие концентрации кислорода и процессы оксигенации – деоксигенации гемоглобина обуславливают образование высокореакционных интермедиатов кислорода, вызывающих нарушение нормального функционирования клетки. Существует антиоксидантная система защиты клетки от свободнорадикального окисления. В её состав входит ряд ферментов и небелковых веществ. Важную роль в антиоксидантной системе играет вещество небелковой природы – аскорбат. Он обладает широким спектром антиоксидантных свойств, в частности, только аскорбат достаточно реакционноспособен для эффективного ингибирования инициации перекисного окисления липидов. Аскорбат блокирует поглощение кислорода и образование перекиси водорода; присутствие аскорбата в клетках оказывает защитное действие на гемоглобин, препятствуя его окислению. Аскорбат в ходе выполнения своих биохимических функций обратимо переходит в окисленную форму – ДАК и ДКГК. Основную роль в биохимических процессах играет редокс-пара – АК/ДАК. По данным литературы, это соотношение может меняться при различных патологических процессах, одним из наиболее распространённых из них является инсулинзависимый сахарный диабет. Исследования, направленные на изучение изменения содержания АК, ДАК и ДКГК в клетках могут быть одним из критериев, свидетельствующих о наличии в организме вышеуказанных процессов.
Целью данной работы явилось определение содержания АК, ДАК, ДКГК в общей эритроцитарной массе у детей, страдающих инсулинзависимым сахарным диабетом. Данная работа представляет собой часть исследований, проводимых на кафедре биохимии и физиологии человека и животных КГУ по изучению метаболизма эритроцитов.
ГЛАВА 1. ОБЗОР ЛИТЕРАТУРЫ 1.1.биохимические процессы при созревании и старении эритроцитов 1.1.1. Характеристика эритроцитов
Зрелый эритроцит человека является упрощенной клеткой по биохимической и структурной организации. Это высокоспециализированная безъядерная клетка. Эритроциты человека образуются из ядросодержащих клеток преимущественно в костном мозге. В этих предшественниках эритроцитов содержатся субклеточные структуры и ферментные системы, необходимые для деления, созревания, дифференцировки, процессов биосинтеза ДНК, РНК, белков, в том числе глобина, синтеза гема, липидов, углеводов, других соединений. На этой стадии развития эритроцита осуществляются окислительные процессы, тканевое дыхание, анаэробное расщепление углеводов (гликолиз), прямое окисление глюкозы через пентозофосфатный путь (Черняк Н.Б., 1976).
До сих пор нет достаточно чётких представлений о том, как соотносятся отдельные стадии созревания ядерных клеток с изменениями химического состава и обмена веществ. Однако известно, что в процессе развития клетки на стадии нормобласта уменьшается количество РНК, увеличивается содержание гемоглобина и утрачивается способность к синтезу ДНК, в связи с чем нарушается способность к митотическому делению.
Ретикулоциты – безъядерные клетки, образующиеся на последнем этапе созревания, предшествующем образованию эритроцитов, характеризуются схожей морфологией, в частности, содержат митохондрии, рибосомы, ЭПР. В ретикулоцитах осуществляется биосинтез глобина, гема, пуринов, пиридиннуклеотидов, фосфатидов, липидов (Фёдоров Н.А., Черняк Н.Б., 1976). РНК практически не синтезируется. Происходит фосфорилирование, сопряжённое с окислением, и гликолиз (Гинодман Л.М., 1968). В обмене веществ ретикулоцитов участвуют эндогенные и экзогенные субстраты, в том числе аминокислоты, глюкоза.
Последний этап созревания – превращение ретикулоцита в эритроцит – протекает 1-3 дня. Происходят значительные изменения в обмене веществ и морфологии клеток (Фёдоров Н.А., 1976).
В зрелых безъядерных эритроцитах нарушены биологический аппарат дыхания, системы синтеза белка, пуринов, порфиринов. Сохраняется способность к гликолизу, утилизации небольшого количества глюкозы в пентозном цикле и синтезу некоторых соединений, например, глутатиона.
В норме длительность жизни эритроцитов поддерживается в течение 120 дней специализированными ферментными системами. Выведение эритроцитов из циркуляции связано с изменениями (структурных компонентов, химического состава, источников энергии), характеризующими старение клеток. Наиболее характерными изменениями при старении эритроцитов являются:
1) уменьшение активности различных ферментов гликолиза и пентозного цикла, что понижает интенсивность данных процессов (Мортенсен, Брайн, 1974);
2) уменьшение содержания липидов, что приводит к изменению структуры эритроцитов, увеличению чувствительности к осмотическому лизису и механическим воздействиям;
3) изменения в составе катионов в результате изменения проницаемости мембраны;
4) изменение содержания АТР, что в свою очередь связывается как с одной из причин нарушения проницаемости, так и с уменьшением приживаемости эритроцитов в кровяном русле.
Одной из ведущих гипотез старения является свободнорадикальная гипотеза, предложенная Д. Хартманом. Она связывает причины возрастных изменений с накоплением молекулярных повреждений в мембранах и генетическом аппарате клетки свободными радикалами и продуктами перекисного окисления липидов. Нарушение нормального функционирования клетки обусловлено высокими скоростями образования высокореакционных интермедиатов кислорода (супероксидрадикал, пероксид водорода, гидроксильный радикал), что, в свою очередь, связано с постоянно протекающими процессами оксигенации и деоксигенации гемоглобина и наличием высоких концентраций кислорода в ходе выполнения основной функции эритроцитов – транспорта кислорода от клетки к тканям и углекислого газа в обратном направлении.
1.1.2. Энергетический обмен в эритроцитахДля поддержания функциональной активности клеток организма необходима затрата энергии. Зрелые эритроциты, циркулирующие в кровяном русле, являются метаболически активными клетками, несмотря на отсутствие способности к синтезу белков, аэробному расщеплению глюкозы в лимоннокислом цикле Кребса (Владимиров Г.Е. по Рапопорту, 1970). Основным процессом обмена энергии в них является гликолиз. Процесс, протекающий в эритроцитах, близок к процессам в других клетках и тканях, и подробно описан (Фёдоров Н.А. по Райкеру, 1976).
К особенностям гликолиза в эритроцитах можно отнести использование, помимо глюкозы, других моносахаридов: фруктозы, маннозы, галактозы, а также инозина, сорбита при наличии соответствующих ферментов (Йошикава, 1968). В процессе гликолиза происходит образование АТР и NADH. Энергия гликолиза используется для активного транспорта катионов через клеточную мембрану и поддержания соотношения между ионами калия и натрия в эритроцитах и плазме, для сохранения целостности мембраны и двояковогнутой формы клетки. Образующийся NADH используется для восстановления пировиноградной кислоты в молочную и для восстановления метгемоглобина при участии метгемоглобинредуктазы. В составе метгемоглобина содержится трёхвалентное железо, вследствие чего он не способен к транспорту кислорода. Характерной особенностью гликолиза в эритроцитах является превращение 1,3- дифосфоглицерата не только в 3-фосфоглицерат, но и в 2,3-дифосфоглицериновую кислоту под действием дифосфоглицеромутазы. 2,3-дифосфоглицерат имеет, наряду с АТР, важное значение в регуляции сродства гемоглобина к кислороду. По мере старения эритроцита происходит уменьшение способности к восстановлению метгемоглобина в гемоглобин, т.е. нарушение функциональной активности эритроцита. Это связанно именно с уменьшением интенсивности гликолиза, в результате которого образуется NADH, необходимый для действия метгемоглобинредуктазы. Уменьшение содержания 2,3-дифосфоглицерата приводит к сдвигу диссоциационной кривой влево, ухудшению отдачи кислорода тканям.
Итогом всех реакций гликолиза является превращение 1 молекулы глюкозы в 2 молекулы молочной кислоты с одновременным превращением 2 молекул ADP в 2 молекулы АТР.
Наряду с гликолизом – анаэробным расщеплением глюкозы до молочной кислоты – в эритроцитах существует дополнительный путь утилизации глюкозы – прямое окисление до углекислого газа и воды в ходе пентозофосфатного цикла. Этот путь неотличим от подобных процессов, протекающих в других клетках и тканях; суммарным результатом цикла является окисление одной из 6 молекул глюкозо-6-фосфата до 6 молекул СО2 и восстановление 12 молекул NADPH. Роль пентозного цикла в зрелых эритроцитах заключается, с одной стороны, в образовании пентозофосфатов. В реакции цикла образуется 3-глицероальдегидфосфат, подвергающийся превращениям в цепи гликолитических реакций и, таким образом, является дополнительным источником энергии. Основное значение пентозофосфатного цикла заключено в образовании молекул NADPH. Значение NADPH определяется его участием в ряде реакций, необходимых для поддержания функциональной активности и целостности эритроцитов. К ним относятся восстановление метгемоглобина в гемоглобин при участии NADPH и метгемоглобинредуктазы и восстановление окисленного глутатиона с помощь. NADPH- глутатионредуктазы. Восстановленный глутатион (GSH), форма со свободно реагирующей тиоловой группой составляет в эритроцитах до 96% общего количества. Сохранение глутатиона в восстановленном состоянии необходимо для предохранения ряда ферментов, содержащих SH- группы, от инактивации, ограждение мембраны клетки от действия перекисей и необратимого окислительного денатурирования гемоглобина.
1.1.3. Антиоксидантная система эритроцитовОсновная функция эритроцитов – транспорт кислорода от лёгких к тканям и СО2 в обратном направлении. Благодаря высоким концентрациям кислорода и постоянно протекающим процессам оксигенации – деоксигенации гемоглобина, в этих клетках с высокой скоростью идут процессы образования свободных радикалов: Н2О2, ОН-. Кроме того, в эритроцитах в результате аутокаталитических реакций образуются перекиси и гидроперекиси липидов.
Основное количество О2- в эритроцитах образуется при аутоокислении гемоглобина в метгемоглобин. Это пример генерации супероксидного радикала, связанной с неферментативным окислением субстрата:
Hb + O2 Û Hb…O2 Û MetHb + O2-
Большую роль в защите клетки от свободных радикалов играют ферментативные антиоксиданты. Эритроциты содержат высокоактивную супероксиддисмутазу, которая осуществляет дисмутацию двух O2- с образованием перекиси водорода:
O2- + O2- Û H2O2 + O2
Образовавшаяся перекись водорода, являющаяся сильнейшим окислителем, частично нейтрализуется неферментативным путём при непосредственном участии аскорбата или других антиоксидантов (a-токоферол, глутатион восстановленный). Основное количество Н2О2 расщепляется в реакциях, катализируемых каталазой и глутатионпероксидазой:
Н2О2 + Н2О2 Þ 2Н2О + О2
Н2О2 + RH2 Þ 2Н2О + R
Важную роль в антиоксидантной системе эритроцитов играют легкоокисляющиеся пептиды, содержащие аминокислоты с SH-группой: метионин, цистеин. Особое место занимает глутатион – трипептид, образованный цистеином, глутаматом, глицином. В организме он присутствует в окисленной и восстановленной форме (GSH). Основной антиоксидантный эффект глутатион оказывает, участвуя в работе ферментативных антиоксидантов. Глутатион является ингибитором активированных кислородных радикалов и стабилизатором мембран. Это связано с тем, что SH- содержащие соединения подвергаются окислению в первую очередь, что предохраняет от окисления другие функциональные группы.
Немаловажный вклад в защиту клетки от органических радикалов вносят неферментативные антиоксиданты. Эффективными перехватчиками органических радикалов являются фенольные антиоксиданты, имеющие в структуре ароматическое кольцо, связанное с одной или несколькими гидроксильными группами. Имеется несколько тысяч фенольных соединений, обладающих антиоксидантным эффектом: витамины группы Е и К, триптофан, фенилаланин, убихиноны, большинство животных и растительных (каротиноиды, флавоноиды) пигментов. Синтезируется ароматическое кольцо только у высших растений и микроорганизмов, поэтому многие из фенольных антиоксидантов входят в группу облигатных пищевых, которые эффективно ингибируют О2- , ОН- и индуцируемые ими процессы перекисного окисления (Оксенгендлер, 1985).
Антиоксидантными свойствами обладают хелатные соединения, связывающие металлы переменной валентности (церулоплазмин, мочевая кислота, трансферрин). Тем самым они препятствуют вовлечению их в реакции разложения перекисей, поскольку в присутствии металлов переменной валентности образование высокореакционных радикалов усиливается (Эристер, 1987).
Таким образом, развитие и функционирование клеток в кислородсодержащей среде не представляется возможным без существования защитных систем – специализированных ферментативных и неферментативных антиоксидантов. В живых организмах постоянен процесс образования прооксидантов, уравновешиваемый дезактивацией их антиоксидантными системами. Для поддержания гомеостаза регенерация антиоксидантов должна быть непрерывной. Отсутствие или нарушение в её непрерывной работе приводит к развитию окислительных процессов, к накоплению окислительных повреждений, что сопровождает ряд патологических физиологических процессов, например, старение (Оксенгендлер, 1985).
1.2. Аскорбат как компонент АОС эритроцитов 1.2.1. Строение и физико-химические свойства аскорбатаВитамин С (L-аскорбиновая кислота) входит в состав алифатического ряда витаминов. По своему строению он может быть отнесен к производным углеводов. Это γ-лактон 2,3-дегидро-L-гулоновой кислоты, производное ненасыщенных полиокси-γ-лактонов. Структура близка структуре a-глюкозы.
Благодаря наличию двух асимметричных атомов углерода в 4 и 5 положениях, аскорбиновая кислота (АК) образует 4 оптических изомера и 2 рацемата. D- и L- аскорбиновые кислоты в природе не встречаются и синтезированы искусственным путём.
Наличие в АК двух сопряжённых двойных связей (углерод-углеродной и углерод-кислородной) обуславливает ее способность к обратимому окислению, продуктом которого является дегидроаскорбиновая кислота (ДАК). ДАК устойчива, но ее лактонное кольцо, в отличие от стабилизированного двойной связью лактонного кольца L-АК в водном растворе легко гидролизуется с образованием 2,3-дикетогулоновой кислоты (2,3-ДКГК). Эта реакция необратима, ее скорость возрастает при повышении температуры и рН среды. Через ряд дальнейших превращений ДКГК переходит в щавелевую и L-треоновую кислоты. Такое же превраще
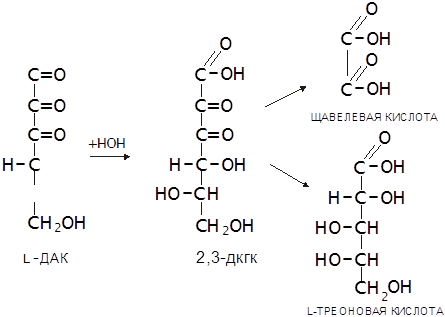 |
ние имеет место в организме (Халмурадов, Тоцкий, 1993):
Способность к О-В превращениям, связанная с ендольной группировкой, которая стабилизирована находящейся в цикле соседней карбонильной группировкой, сопровождающаяся перенесением атомов водорода к акцепторам, является важнейшей каталитической функцией АК в живом организме. L-АК по своей биологической активности высокоспецифична. Витаминная активность проявляется только при наличии свободных гидроксильных групп. Различные функциональные производные по ним лишают молекулу витаминной активности почти полностью, как и гидрирование ненасыщенной связи лактонного кольца. Поэтому L-ДАК имеет витаминную активность, равноценную L-АК, тогда как 2,3-ДКГК полностью ее лишена. Вследствие легкой окисляемости L-АК – донор Н+, она количественно легко восстанавливает многочисленные соединения, как-то: йод, перманганат калия и другие. L-АК – переносчик Н+ в некоторых ферментативных реакциях живой клетки, она легко окисляется пероксидазой, цитохромоксидазой, каталазой. L-АК восстанавливает окисленные формы ферментов, окисляясь в ДАК, обратимо легко регенерирующуюся в АК под действием глутатиона за счет его сульфгидрильной группы:
![]()
![]()
![]()
![]()
Окисление АК катализируется медью, в меньшей степени – катионами серебра и железа. Имеется предположение, что специфическим катализатором окисления АК в животных организмах является белок, синтезирующийся в печени, осуществляющий транспорт меди, обладающий оксигеназной активностью, - церулоплазмин. В меньшей степени окисление аскорбата катализируют другие катионы, в частности, серебра и железа. Комплексоны, флавоноиды тормозят окислительный распад АК. Некоторые белки ингибируют окисление АК, связываясь с ней или путём образования комплекса с медью – сывороточные глобулины (Борец, 1980). Окисление тормозится –SH содержащими соединениями: сернистая кислота блокирует фермент аскорбиназу; С-SH связывает ионы Cu+, удаляя т. с. катализатор окисления АК из реакции (Киверин, 1971).
0 комментариев