Навигация
Образование ацетилкофермента А, в котором принимает участие ацетат, кофермент А и АТФ. Реакция катализируется ацилкофермент А-синтетазой
15586
знаков
0
таблиц
10
изображений
1. Образование ацетилкофермента А, в котором принимает участие ацетат, кофермент А и АТФ. Реакция катализируется ацилкофермент А-синтетазой.
![]()
2. Образование сукцинилкофермента А из двух молекул ацетилкофермента А. Считается не менее вероятным другой путь: вовлечение ацетата в цикл Кребса и образование в нём сукцината и затем сукцинилкофермента А. Некоторые исследователи считают исходным субстратом биосинтеза хлорофилла именно сукцинилкофермент А, не рассматривая реакции его образования (как не специфичные, осуществляющиеся в связи и с другими метаболическими цепочками).
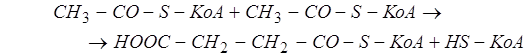
3. Образование ![]() -амино-
-амино-![]() -кетоадипиновой кислоты из сукцинилкофермента А и глицина, катализируемого так же, как и следующая реакция, ферментом синтетазой
-кетоадипиновой кислоты из сукцинилкофермента А и глицина, катализируемого так же, как и следующая реакция, ферментом синтетазой ![]() -аминолевулиновой кислоты:
-аминолевулиновой кислоты:
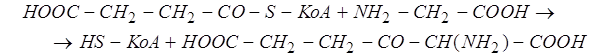
4. Из ![]() -амино-
-амино-![]() -кетоадипиновой кислоты путём декарбоксилирования образуется
-кетоадипиновой кислоты путём декарбоксилирования образуется ![]() -аминолевулиновая кислота:
-аминолевулиновая кислота:
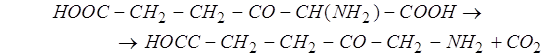
5. Синтез из двух молекул ![]() -аминолевулиновой кислоты пирроленинового кольца и затем изомеризация его в пиррольное кольцо с образованием порфобилиногена. Реакция катализируется ферментом дегидразой
-аминолевулиновой кислоты пирроленинового кольца и затем изомеризация его в пиррольное кольцо с образованием порфобилиногена. Реакция катализируется ферментом дегидразой ![]() -аминолевулиновой кислоты.
-аминолевулиновой кислоты.
Второй этап включает реакции синтеза из четырёх пиррольных колец одной молекулы протопорфирина.
6. Из четырех молекул порфобилиногема под влиянием фермента порфобилиноге-дезаминазы синтезируется цепочка тетрапиррана.
7. Наименее изучен механизм реакции замыкания открытой цепи тетрапиррана в уропорфириноген III.
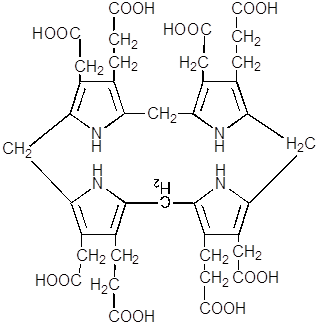
8. В результате декарбоксилирования всех четырёх остатков ацетата из уропорфириногена III образуется копропорфириноген III, фермент – уропорфириногендекарбоксилаза.
9. Происходит декарбоксилирование и дегидрирование двух из четырёх пропионатных остатков, что приводит к появлению винильных радикалов в кольцах и образованию протопорфириногена IX, фермент – копропорфириногендекарбоксилаза.
10.
 |
В результате дегидрирования протопорфириногена IX появляется протопорфирин IX.
Третий этап характеризуется образованием и превращением магнийпорфиринов.
11. Протопорфирин, взаимодействуя с магнием, превращается в магнийпротопорфирин.
12.
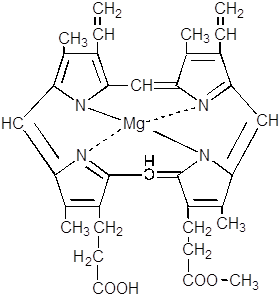 |
Далее следует ферментативная этерификация метилом пропионатного остатка и образование монометилового эфира магний протопорфирина.
13.
 |
Последний превращается в протохлорофиллид в серии реакций, включающих преобразование метильного производного пропионата.
14.
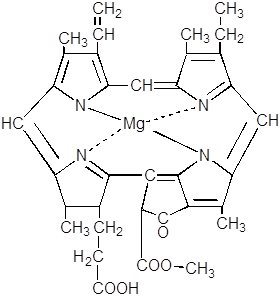 |
Превращение протохлорофиллида в хлорофиллид, заключающееся в гидрировании одной из двух двойных связей кольца, осуществляется, как правило, при участии света (в ходе фотохимической реакции).
Лишь у некоторых низших и голосеменных растений эта реакция может протекать ферментативно в темноте. В хлорофиллид превращается не свободная форма протохлорофиллида, а связанная с белком в единый комплекс – так называемый протохлорофиллидголохром.
15. Последняя реакция – ферментативная этерификация хлорофиллида фитолом, в результате чего образуется хлорофилл а.
Поскольку синтез хлорофилла – многоэтапный, в нём участвуют различные ферменты, составляющие, по-видимому, полиферментный комплекс. Интересно заметить, что образование многих из этих белков-ферментов ускоряется на свету. Содержание хлорофилла в листе колеблется незначительно. Это связано с тем, что идёт непрерывный процесс разрушения старых молекул и образование новых молекул хлорофилла. Причём эти два процесса уравновешивают друг друга. При этом предполагается, что вновь образовавшиеся молекулы хлорофилла не смешиваются со старыми и имеют несколько иные свойства.
Различные виды хлорофилла.
Различные виды хлорофилла отличаются, как правило, природой заместителей при ![]() -атомах углерода в пиррольных кольцах порфиринов. Все связи
-атомах углерода в пиррольных кольцах порфиринов. Все связи ![]() -углеродных атомов заняты в формировании порфиринового кольца и поэтому не могут определять специфику отдельных видов хлорофилла. Хлорофиллы бактерий называют бактериохлорофиллами. Их известно четыре. Большинство пурпурных бактерий содержит бактериохлорофилл а, который и обуславливает их способность к фотосинтезу:
-углеродных атомов заняты в формировании порфиринового кольца и поэтому не могут определять специфику отдельных видов хлорофилла. Хлорофиллы бактерий называют бактериохлорофиллами. Их известно четыре. Большинство пурпурных бактерий содержит бактериохлорофилл а, который и обуславливает их способность к фотосинтезу:
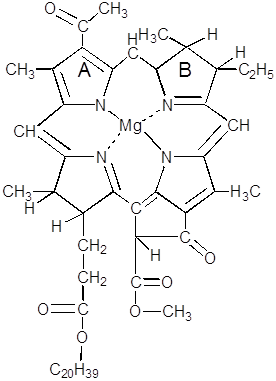
Этот пигмент представляет собой порфирин, у которого ![]() -углеродные атомы имеют следующих заместителей в положениях: 1 – метил, 2 – ацетил, 3 – метил, 4 – этил, 5 – метил, 6-й атом углерода участвует в образовании насыщенного циклопентанового кольца, 7 – сложный эфир пропионовой кислоты и высокомолекулярного ненасыщенного спирта фитола, 8 – метил. Кольца В и D имеют лишь по одной двойной связи. В циклопентановом кольце в положении 9 – кетогруппа, а в 10 – карбоксиметильный радикал
-углеродные атомы имеют следующих заместителей в положениях: 1 – метил, 2 – ацетил, 3 – метил, 4 – этил, 5 – метил, 6-й атом углерода участвует в образовании насыщенного циклопентанового кольца, 7 – сложный эфир пропионовой кислоты и высокомолекулярного ненасыщенного спирта фитола, 8 – метил. Кольца В и D имеют лишь по одной двойной связи. В циклопентановом кольце в положении 9 – кетогруппа, а в 10 – карбоксиметильный радикал ![]() . Фитол может рассматриваться как производное изопрена или дитерпена с одной двойной связью.
. Фитол может рассматриваться как производное изопрена или дитерпена с одной двойной связью.
Бактериохлорофиллы с и d зелёных бактерий отличаются от бактериохлорофилла а рядом особенностей. Они не имеют циклопентановного кольца. Карбоксиметильный радикал при атоме 10 отсутствует, а атомы 9 и 10 входят в состав пропила. Вместо фитольного остатка содержат фарнезильный -- ![]() .
.
При втором углеродном атоме порфирин имеется вместо ацетильного оксиэтильный радикал ![]() . У некоторых зелёных бактерий имеется несколько процентов (до 10) бактериохлорофилла а.
. У некоторых зелёных бактерий имеется несколько процентов (до 10) бактериохлорофилла а.
Все другие фотосинтезирующие организмы содержат в качестве основного зелёного пигмента хлорофилл а.
От бактериохлорофилла а он отличается тем, что в положении 2 имеет винильный радикал ![]() , а кольцо В имеет ещё одну двойную связь (за счёт того, что содержит на 2 атома водорода меньше).
, а кольцо В имеет ещё одну двойную связь (за счёт того, что содержит на 2 атома водорода меньше).
Кроме хлорофилла а, все высшие растения и большинство водорослей (за исключением сине-зелёных и красных) содержат хлорофилл. Он отличается от хлорофилла а тем, что при углеродном атоме 3 вместо метильной имеется формильная группа СНО.
У бурых и хризофитовых водорослей, а также динофлагеллят обнаружен хлорофилл с, не имеющий остатка фитола.
Красные водоросли имеют хлорофилл, который отличается от хлорофилла а тем, что вместо винильной группы при углеродном атоме 2 имеется формильный радикал.
Распознавание различных видов хлорофилла осуществляется с помощью спектральных характеристик. Обычно изучают спектр поглощения растворов пигментов, реже – спектр люминесценции.
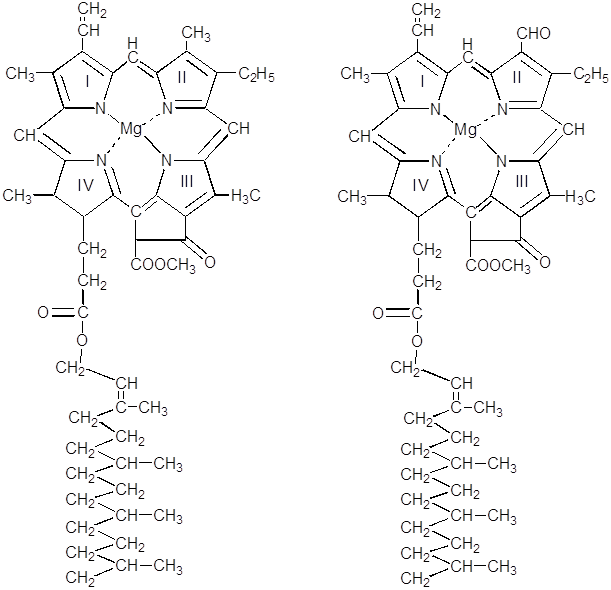
хлорофилл а хлорофилл b
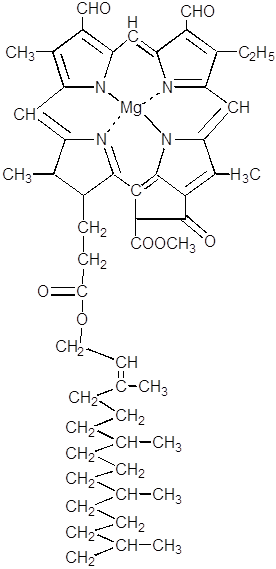
Хлорофилл d
Хлорофилл а

Хлорофилл b
Министерство высшего и профессионального образования Российской Федерации
Тверской Государственный Технический Университет
Кафедра информатики и прикладной математики
Курсовая работа«Хлорофилл: его свойства и биосинтез»
Выполнил: студентка группы БТ-111
Николаева Е. С.
Проверил: Семёнов А. В.
Принял: Семёнов А. В.
Тверь, 2001Похожие работы
... способной координировать ионы металлов М2+, М3+, М4+ и даже с большей степенью окисления. В результате комплексообразования образуются комплексные соединения порфиринов, т.н. металлопорфирины, обладающие многообразными структурными и химическими особенностями, высокой биологической и каталитической активностью. При этом металл либо занимает центр полости N4 и оказывается в экваториальной плоскости ...
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... как в результате их жизнедеятельности азот, находящийся в виде соединений, недоступных для усвоения растениями, превращается в соли азотной кислоты, которые хорошо ими усваиваются. 2. Биосинтез белков Любая живая клетка способна синтезировать белки, и эта способность представляет одно из наиболее важных и характерных ее свойств. С особенной энергией идет биосинтез белков в период роста и ...
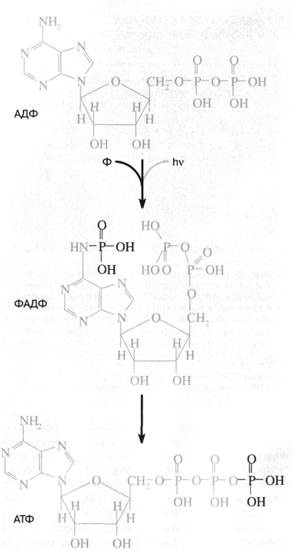

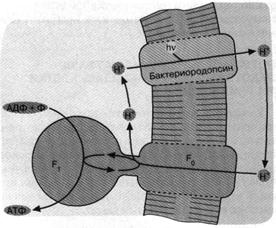
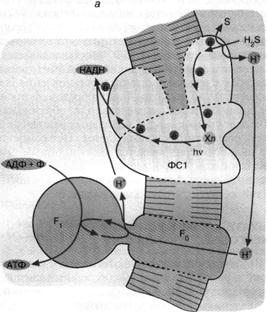
Перенос фосфорила с аде-ниновой "головы" нуклеотида на фосфатный "хвост" должен сопровождаться его стабилизацией, поскольку весьма лабильный фосфоамид заменяется на менее лабильный фосфоангидрид (рис. 1). Стадии 2 и 3 гипотетичны и призваны объяснить механизм синтеза АТФ под действием ультрафиолетового света в опытах С. Понамперумы и сотрудников [3]. АДЕНИНСОДЕРЖАЩИЕ КОФЕРМЕНТЫ Аденин ...
0 комментариев