Сравнительная характеристика уровня элементарной рассудочной деятельности (элементарного мышления) у животных разных таксономических групп
Рассудочная деятельность и сложность строения мозга
Сопоставление способности к экстраполяции и к обучению. Роль экологических факторов в успешности решения тестов
Элементарное мышление человекообразных и низших обезьян
Навигация
Рассудочная деятельность и сложность строения мозга
Сравнительная характеристика и морфофизиологические основы мышления животных
43638
знаков
0
таблиц
1
изображение
3. Рассудочная деятельность и сложность строения мозга.
Л.В. Крушинский и его сотрудники в 70-е годы XX века предприняли исследование морфофизиологических основ элементарной рассудочной деятельности животных. Эти работы включали сопоставление сложности строения высших ассоциативных структур переднего мозга птиц и млекопитающих с уровнем развития у них способности к рассудочной деятельности. На основе таких сопоставлений была изучена роль отдельных образований мозга в способности к экстраполяции и в способности к обучению. Ряд работ был специально посвящен сравнению способности животных к решению элементарных логических задач и к обучению.
3.1. «Цефализация», сложность нейронного строения и уровень рассудочной деятельности.
В лаборатории Л. В. Крушинского изучалась связь сложности организации мозга и общего уровня эволюционного развития в классах птиц и млекопитающих. Нейроморфологические данные, накопленные к 70-м годам XX в., свидетельствовали, что индекс цефализа-ции — относительный объем высших отделов мозга (новой коры у млекопитающих и гипер- и неостриатума у птиц) растет по мере повышения уровня эволюционного развития вида. Л. В. Крушинский (1986) показал, что как у птиц, так и у млекопитающих степень цефализации в пределах каждого класса позвоночных представляет собой существенный параметр, определяющий уровень развития рассудочной деятельности.
А. Портман (Portmann, I946) получил следующие величины индексов относительного объема полушарий птиц: голубь — 4,0; курица — 3,27; утка (Anas platirhinchos) — 6,08; сокол (Falco tinninculus) — 8,24; канюк (Buteo buteo) — 9,78; сорока (Pica pica) — 15,81; грач (Corvus frugilegus) — 15,68; ворона (ы) — 15,3
У млекопитающих также обнаруживается зависимость между уровнем развития элементарной рассудочной деятельности и относительным размером мозга. Л. В. Крушинский приводит следующие величины квадратического показателя головного мозга (по Я. Я. Рогинскому) для ряда видов млекопитающих: мышь — 0,0088; крыса — 0,0123; кролик — 0,0705; кошка — 0,195; собака — 0,464. Приматы и дельфины обладают наиболее дифференцированным и крупным мозгом среди млекопитающих.
В ряду млекопитающих происходит увеличение площади ассоциативных зон коры больших полушарий, в частности префронтальной (лобной) области. Это тоже является индикатором усложнения строения высших отделов мозга. Такая же закономерность описана и в отношении ассоциативных областей мозга птиц.
В этой связи следует отметить и еще одно немаловажное обстоятельство. Сравнительные исследования Л. В. Крушинского и его сотрудников (1986) показали, что нет прямой и непременной связи между степенью развития элементарной рассудочной деятельности и наличием новой коры. Мозг птиц построен по иному плану, чем мозг млекопитающих. В процессе филогенеза особого развития у них достигли новые, отсутствующие у млекопитающих, отделы стриатума (гипер- и неостриатум— wulst), в то время как у млекопитающих сформировалась новая кора. Именно за счет этих отделов стриатума увеличился объем полушарий и переднего мозга врановых птиц
Наряду с увеличением относительных размеров высших отделов мозга, другим важным фактором, определяющим степень развития рассудочной деятельности, следует считать сложность нейронной организации мозга. В филогенетическом ряду позвоночных наблюдается увеличение разнообразия нейронного строения мозга, с прогрессирующим увеличением числа так называемых звездчатых нейронов (Богословская, Поляков, 1981), а также усложнение систем контактов между нейронами. Были выявлены особенности цитоархитектони-ки переднего мозга вороны и голубя (Крушинский и др., 1985; Воронов, 1996).
Особенно сложным строением отличаются нейроны гиперстриа-тума вороны. Они имеют более длинные и более извилистые дендри-ты, что определяет большее число контактов с другими клетками. Количество шипиков на дендритах также значительно больше, чем на дендритах нейронов мозга голубя.
Специфическая особенность строения мозга птиц — так называмые мультинейронные комплексы (Богословская, Поляков, 1981). Это клеточные ассоциации сложной пространственной структуры, состоящие, по-видимому, из функционально связанных нервных элементов. Исследования Д. К. Обухова (Обухов, 1996; Андреева, Обухов, 1999) демонстрируют конкретное строение таких мультинейронных комплексов: у ворон они могут включать до 20 нейронов, у голубей до 10.
В двух классах позвоночных — млекопитающих и птиц — усложнение строения мозга в ряду видов коррелирует с повышением уровня развития их элементарного мышления. Это прослеживается и в увеличении числа тестов, которые более развитые животные способны решать, и в повышении уровня сложности этих тестов.
3.2. Повреждение участков мозга и способность к экстраполяции.
Анализ морфофизиологических основ рассудочной деятельности проводился в 60—70-е годы в лаборатории Л. В. Крушинского. В опытах Д. А. Флесса (1974) был начат фармакологический анализ способности к экстраполяции. Было показано, что введением холинолитиков она блокировалась, но сохранялась при использовании препаратов, выключающих катехоламинергическую нейромедиаторную систему мозга (даже при использовании очень высоких дозировок).
Более подробно была исследована роль различных структур мозга у животных, обладающих разной способностью к экстраполяции, в опытах с их повреждением (удалением, экстирпацией). Для анализа последствий разрушений участков мозга были выбраны представители разных классов, в пределах класса — разные виды, в пределах одного вида — разные группы животных. Часть экспериментов проводили на животных, взятых в опыт впервые, а часть — на животных, имевших до операции опыт решения задачи. Разрушались структуры, ответственные за высшие ассоциативные функции и имеющие тенденцию к развитию и усложнению в пределах своего класса: префронтальная кора млекопитающих, комплекс ядер переднего мозга птиц — Wulst и дорсовент-ральная область коры — у пресмыкающихся, поскольку степень их развития коррелирует с уровнем способности к экстраполяции. В «контрольных» опытах был проанализирован эффект равных по объему разрушений других структур переднего мозга, выполняющих более элементарные функции. Эксперименты проводились на собаках (успешно решающих тест на экстраполяцию), кошках (чьи показатели были ниже), воронах (имеющих высокие показатели во всех тестах), курах (тест не решают), болотных черепахах Emis orbicularis, успешно экстраполирующих (Зорина, Попова, 1978; Зорина, Федотова, 1981; Адрианов, Молодкина и др., 1987; Очинская и др., 1988).
У млекопитающих, решавших задачу впервые, способность к экстраполяции нарушалась при повреждении префронтальной коры (высшей ассоциативной зоны) и не страдала при повреждении других областей коры.
У врановых птиц, которым тест был также предъявлен впервые, аналогичные нарушения наблюдались при повреждении Wulst (комплекс ядер гиперстриатума (Karten, 1991), который считают функциональным аналогом префронтальной коры млекопитающих), и отсутствовали при контрольном разрушении гиппокампа (старой коры).
У рептилий (черепах) способность к экстраполяции нарушалась при экстирпации дорсовентральной области коры (зоны перекрытия сенсорных проекций разных модальностей). В мозге рептилий она выполняет как ассоциативные, а так и проекционные функции. При разрушении чисто проекционной области выполнение теста не нарушалось.
После разрушений участков мозга, имеющих высшие ассоциативные функции, все оперированные животные первое время решали задачу на экстраполяцию чисто случайно. Однако по мере его многократных повторений доля правильных обходов ширмы постепенно увеличивалась, т.е. животное обучалось навыку обхода ширмы со стороны движения корма (см. также ниже).
Если разрушение производили у животных, успешно решавших задачу до операции, способность к экстраполяции в целом сохранялась, хотя доля правильных решений снижалась. Следовательно, разрушение указанных структур не нарушает способности животных к обучению, но практически исключает проявление способности к экстраполяции при первом предъявлении задачи. Сходство эффектов разрушений у представителей всех трех классов отражает, по-видимому, общность процессов, лежащих в основе решения этого теста.
В опытах с птицами в качестве «контрольной» структуры, разрушение которой не должно было изменить уровня решения задачи на экстраполяцию, была выбрана старая кора (гиппокамп). В главе 3 (см. 3.4.3) было показано, что повреждение этой структуры у птиц и млекопитающих существенно ухудшает процесс запоминания. В наших опытах при разрушении гиппокампа поведение ворон (не имевших опыта решения задачи до операции) не изменялось ни в первом, ни при повторных предъявлениях теста на экстраполяцию. У кур (низкий уровень рассудочной деятельности) свойственное им постепенное формирование условно-рефлекторного навыка обхода ширмы после разрушения гиппокампа замедлялось, в то время как при разрушении гиперстриатума их поведение не изменялось.
Сходным образом разрушение хвостатого ядра не ухудшало решения задачи на экстраполяцию у кошек, но ослабило эту способность у крыс (выраженную слабо даже в норме).
У представителей млекопитающих, птиц и рептилий выявлены структуры мозга, в наибольшей степени связанные с осуществлением решения задачи на экстраполяцию. Их разрушение практически исключает проявление способности к экстраполяции при первом предъявлении задачи, ухудшает показатели животных, решавших задачу до операции, но не препятствует обучению при многократных предъявлениях. Различие влияний на рассудочную деятельность и на обучение сходным задачам позволяет говорить об участии в этих процессах разных нейрофизиологических механизмов. Сходство эффектов разрушений у представителей всех трех классов отражает общность процессов, лежащих в основе решения этого теста.
Сопоставление решения задачи на экстраполяцию и обучения сходному навыку проводилось и в других экспериментах.
Похожие работы
... концепцию соотношения врожденного и приобретенного в формировании целостного поведенческого акта (концепция так называемых «унитарных реакций»). Наибольшую известность получили его исследования мышления животных. В конце 50-х годов Л. В. Крушинский совместно с сотрудниками своей лаборатории приступил к многоплановому физио-лого-генетическому исследованию зачатков мышления у широкого диапазона ...

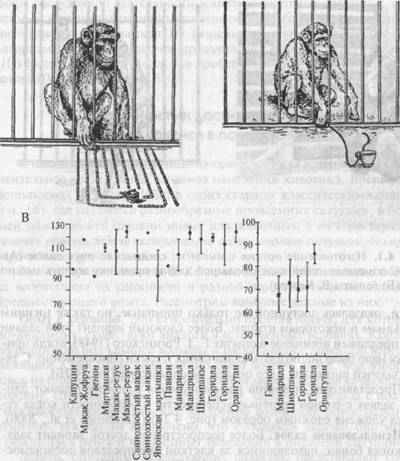



... «Социальное сознание» (Social Cognition) 1. Самоузнавание в зеркале 2. «Перемена ролей» 3. Преднамеренный обман В этой главе приводятся наиболее известные методы изучения тех форм рассудочной деятельности животных, которые связаны с экстренным улавливанием принципа задачи (!) и экстренной реорганизацией независимых навыков (И). Решение на основе выявления общего алгоритма при многократном ...


... важнейших вопросов антропологии, этнографии, демографии, исторической и медицинской географии. Расы - это территориальные группы людей, выделяемые на основе их генетического родства, которое проявляется в определенном физическом сходстве. Большинство отечественных антропологов различают три большие расы человечества - монголоидную ("желтую"), европеоидную ("белую") и негроидно-австролоидную (" ...
... человека эволюционно предшествуют некоторые элементы психического поведения у высших животных. К ним относится психонервная деятельность, направляемая воспроизведением образов предыдущего опыта, основанная на образном поведении животного, когда основным действенным стимулом для запускания какого-либо поведенческого акта становится не сам реальный объективный стимул окружающей среды, а «нейронный» ...
0 комментариев