Гипоталамус, или подбугровая область промежуточного мозга, является высшим центром интеграции и регуляции вегетативных функций организма. Он принимает участие в корреляции различных соматических функций, регуляции работы желудочно-кишечного тракта, сна и бодрствования, водно-солевого, жирового и углеводного обмена, поддержания температуры тела и гомеостаза. Одна из наиболее важных функций гипоталамуса связана с регуляцией деятельности эндокринной системы организма.
Разнообразие функции гипоталамуса обусловлено сложностью его морфологического строения и обилием связей с различными отделами нервной системы, органами чувств, внутренними органами и внутренней средой организма.
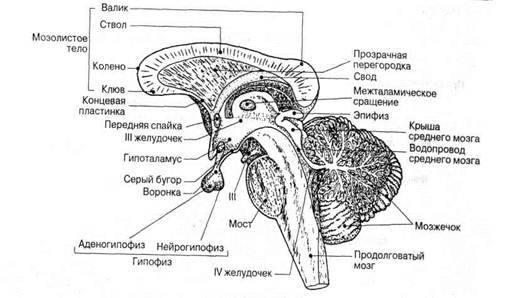
Строение гипоталамуса. Гипоталамус относится к филогенетически древним образованиям мозга и хорошо развит уже у низших позвоночных. Он образует дно третьего желудочка и лежит между перекрестом зрительных нервов и задним краем маммилярных тел. В состав гипоталамуса входит серый бугор, срединное возвышение, воронка и задняя или нервная доля гипофиза. Спереди он граничит с преоптической областью, которую отдельные авторы также включают в систему подбугорья.
Гипоталамус развивается в ранний период эмбриогенеза из переднего мозгового пузыря. В процессе развития головного мозга, после обособления больших полушарий, передний мозговой пузырь дает начало межуточному мозгу, а его полость превращается в третий желудочек. В дне этого желудочка путем выпячивания образуется мозговая воронка, дистальный конец которой превращается в заднюю долю гипофиза. Основание воронки значительно утолщается и дает начало серому бугру. В ка-удальной части образуются парные маммилярные тела. Боковые стенки третьего желудочка образуют зрительные бугры, связанные с большими полушариями головного мозга. Центральное серое вещество гипоталамуса без резкой границы переходит в центральное серое вещество среднего мозга. Нервные клетки в гипоталамусе собраны в более или менее обособленные группы или ядра, которые занимают в нем определенное место и состоят из различных по своему строению невронов. Разнообразие нейрального состава ядер гипоталамуса обусловлено их функциональной дифференциройкой.
В процессе эволюции ряда животных число и структура гипоталамических ядер подверглись значительным изменениям. У кролика, например, насчитывают 30, а у собаки 15 пар ядер.
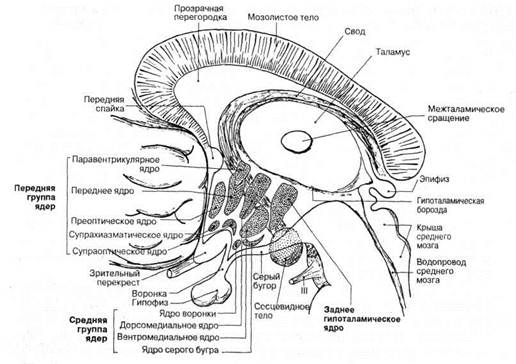
В литературе пока отсутствует единая номенклатура гипоталамических ядер. Пинес и Майман выделяют в гипоталамусе передний, средний и задний отделы. В каждом отделе они различают следующие ядра.
Передний отдел: 1) супрахиазматическое; 2) супраоптическое (передние, латеральные и медиальные отделы); 3) пара-вентрикулярное.
Средний отдел: 1) супраоптическое (задние отделы); 2) ту-беральные (верхние, средние и нижние); 3) паллидо-инфунди-булярное; 4) маммило-инфундибулярное.
Задний отдел: 1) маммило-йнфундибулярные; 2) ядра маммилярных тел (внутреннее, наружное, вставочное); 3) супра-маммилярные.
Филогенетически наиболее древними образованиями гипоталамуса являются паравентрикулярное и супраоптическое ядра. Они гомологичны преоптическим ядрам низших позвоночных. У млекопитающих суираоптическое ядро лежит в переднем гипоталамусе над хиазмой (рис. 1) и проходит в дорсолатеральном направлении от зрительного перекреста до середины серого, бугра.
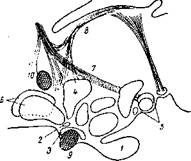 Рис. 72. Сагиттальный срез мозга млекопитающего (по Кларку):
Рис. 72. Сагиттальный срез мозга млекопитающего (по Кларку):
1 — гипофиз; 2 — супрахиазматическое ядро; 3 — супраоптическое ядро; 4 — паравентрикулярное ядро; 5 — ядра мамми-лярной области; 6 — ядра преоптической области; 7 — свод; 8— Stria terminilis — хиазма; 10—передняя комиссура.
У многих животных оно подразделяется на отдельные группы нейронов, соединенных между собой клеточными мостиками. Не менее характерной группировкой клеток гипоталамуса является паравентрикулярное ядро, расположенное под передней коммисурой в стенке третьего желудочка. Паравентрикулярное ядро развивается из того же самого клеточного материала, что и супраоптическое ядро. В строении клеток этих ядер обнаруживается значительное сходство. Они имеют округлую, грушевидную или удлиненную форму и от нейронов других ядер гипоталамуса и центрального серого вещества отличаются значительно более крупными размерами.
 Рис. 2. Схема васкулярных связей гипоталамуса и гипофиза (по Алешину) :
Рис. 2. Схема васкулярных связей гипоталамуса и гипофиза (по Алешину) :
/ — супраоптическое ядро; 2 — паравентрикулярное ядро; 3 — туберальные ядра; 4 — маммилярные ядра; 5 — первичная капиллярная сеть в срединном возвышении; 6 — сосудистые клубочки первичной капиллярной сети; 7 — вены портальной системы; 8 — передняя доля гипофиза; 9 — средняя доля гипофиза; 10 — задняя доля гипофиза; // — вторичная капиллярная система в передней доле гипофиза; 12 — капилляры задней доли гипофиза; 13 — гипоталамо-гипо-физарный тракт.
Васкуляризация гипоталамуса. Гипоталамическая область характеризуется обильным кровоснабжением. Наибольшей васкуляризацией отличается паравентрикулярное и супраоптическое ядра, в которых каждая клетка связана с 2—3 капиллярами. Здесь на площадь 1 мм2 приходится до 2650 капилляров. Электронно-микроскопические исследования показали, что в местах контакта между телом нейрона и сильно утонченной базальной мембраной эндотелия капилляров часто совсем нет глиальной прослойки. Вследствие этого сосуды обладают очень хорошей проницаемостью даже для высокомолекулярных белковых соединений. Из крови в клетки супраоптических и паравентрикулярных ядер легко поступают питательные вещества, гормоны и другие химические соединения. Гипоталамические образования поэтому обладают высокой чувствительностью к отклонениям в составе гуморальной среды организма и отвечают на них изменением физиологической активности.
Важнейшее значение в механизме гипоталамической регуляции гормональной функции гипофиза имеет общность их васкуляризации. Между гипоталамусом и передней долей гипофиза существует специальная система кровообращения, получившая название воротной, или портальной, системы сосудов гипофиза. Она состоит из артериол, которые берут начало от артерий виллизиевого круга. Артериолы проникают в срединное возвышение серого бугра и здесь распадаются на большое количество капилляров. В срединном возвышении клубочки и петли этих капилляров вступают в тесный контакт с окончаниями нервных волокон клеток нейросекреторных ядер гипоталамуса и образуют с ними так называемые вазоневральные синапсы (рис. 2). Первичные капилляры в сером бугре собираются в портальные вены, которые по гипофизарной ножке идут в переднюю долю гипофиза, и в его паренхиме распадаются в густую сеть синуооидных капилляров (вторичная капиллярная сеть). В заднюю долю гипофиза сосуды портальной системы не проникают, и кровь в нее поступает из других источников. Движение крови по портальной системе от гипоталамуса к гипофизу происходит в результате сокращения стенок сосудов. В переднюю долю гипофиза кровь поступает еще по средней и задней гипофизарным артериям, а также по сосудистым анастомозам из нейрогипофиза.
Связи гипоталамуса. Гипоталамическая область имеет обширные связи с различными отделами центральной нервной системы, в том числе с ретикулярной формацией ствола мозга, гипофизом и т. д. Среди проводящих путей гипоталамуса различают эфферентные, афферентные и внутригипоталамические связи.
Эфферентные пути от гипоталамуса идут к таламусу (гипо-таламо-таламический путь), покрышке (маммило-тегментальный путь), от всех ядер гипоталамуса к нижележащим симпатическим образованиям и узлам (диффузные нисходящие связи), от гипоталамуса к гипофизу (гипоталамо-гипофизарный тракт). В гипоталамо-гипофизарных путях выделяют супраоптикогипо-физарный, а также туберо-гипофизарный пути. Первый путь образован большим количеством (до 100000) аксонов клеток су-праоптического и паравентрикулярного ядер, которые по ножке гипофиза поступают в заднюю долю гипофиза. Эти волокна проходят во внешнем слое срединного возвышения и не проникают из задней в переднюю долю гипофиза (см. рис. 2).
В регуляции функций эндокринной системы особое значение имеет тубероинфундибулярный путь, который передает эфферен-тацию из гипоталамуса в переднюю долю гипофиза. Волокна этого пути прослеживаются до срединного возвышения, где их окончания с петлями и клубочками первичных капилляров портальной системы образуют рассмотренные выше вазоневральные синапсы.
Афферентные пути к ядрам гипоталамуса идут от таламуса, лобных долей, гиппокампа, зрительного бугра,- амигдалоидного комплекса, миндалин, экстрапирамидной системы и ретикулярной формации ствола мозга.
Ретикулярной формации приписывается большое значение в регуляции функции гипоталамуса и эндокринной системы. Исследованиями Грина, Русселя и др. установлено, что ядра гипоталамуса находятся в тесной анатомической и.функциональной связи с ретикулярной формацией. Последняя образована сложным комплексом невронов различной величины, которые диф-фузно рассеяны в стволе мозга. Для отростков клеток ретикулярной формации характерно наличие большого количества коллатералей, через посредство которых один аксон может вступать в функциональные связи с множеством нервных клеток (до 20 000). Ретикулярная формация, как впервые было установлено Мэгуном и Мурицци, оказывает общее активирующее действие на различные отделы мозга и состоит из восходящей и- нисходящей систем. Волокна восходящей системы от каудальных частей продолговатого мозга, варолиева моста и среднего мозга проецируются на различные участки коры больших полушарий; нисходящие же волокна связывают ретикулярную формацию с системой спинного мозга.' В ретикулярную формацию проецируется огромное количество волокон от ядерных образований ствола мозга, рецепторов внутренних органов, от аппаратов зрения, слуха и проводников чувствительности. Ретикулярная формация отличается высокой чувствительностью к изменениям гуморальной среды организма. На воздействие гормонов и различных химических соединений она быстро отвечает изменением своей физиологической активности.
Гипоталамус получает восходящие волокна преимущественно от ретикулярной формации среднего мозга. Через покрышку среднего мозга и задний отдел гипоталамуса эти волокна достигают серого бугра. На этой морфологической основе осуще-ствляется функциональная связь между ретикулярной формацией, гипоталамусом и эндокринными железами. Ретикулярная формация среднего мозга передает импульсы через гипоталамус к эндокринным железам организма и оказывает активизирующее действие на ядра подбугорья.
Нейросекреторная деятельность гипоталамуса. Нейроны отдельных ядер гипоталамуса проявляют способность к секреторной деятельности (нейрокринии) и вырабатывают особые вещества (нейросекреты), которые играют важную роль в регуляции функции эндокринной системы. Пионерами в изучении гипотала-мической нейросекреции являются Шаррер и Гаупп, которые еще в 1933 г. обнаружили в клетках переднего гипоталамуса гранулы и капельки нейросекрета. Последующими исследованиями было установлено широкое распространение явлений нейросекреции не только у позвоночных, но и беспозвоночных животных.
У рыб нейросекреторная функция характерна для преоптического ядра передней области гипоталамуса. У высших позвоночных и человека эти свойства в наиболее отчетливой форме выражены в клетках паравентрикулярного и супраоптического ядер. Нейросекреция характерна и для нейронов ядер серого бугра, но в них она не получает такого яркого морфологического выражения, как в паравентрикулярном и супраоптическом ядрах.
Нейросекреторные невроны гипоталамуса одновременно сочетают в себе свойства нервных и железистых клеток. Они относятся к мультиполярным невронам с относительно крупным ядром и содержат нейрофибриллы, хорошо развитую систему эргастоплазмы (субстанцию Ниссля) с рибосомами и общие для всех клеток органоиды.
Нейросекреторные процессы у высших животных наиболее полно изучены в клетках супраоптического и паравентрикулярного ядер. Нейросекрет представлен гранулярными образованиями, обладающими известным постоянством своего строения у всех животных. Гранулы имеют вид гомогенных шариков и пузырьков, окруженных мембраной. В зависимости от специфического отношения к красителям различают гомориположительный и гомориотрицательный нейросекрет. Первый хорошо окрашивается хромовоквасцовым гематоксилином по Гомори в темно-синий цвет; гомориотрицательное или оксифильное нейросекре-торное вещество при окраске по Гомори красится флоксином в розовый цвет.
Химическая природа нейросекрета окончательно не выяснена. Гомориположительное вещество обладает относительно высокой химической устойчивостью и является сложным белково-полиса-харидо-липидным соединением. Гомориотрицательный нейросекрет является относительно простым протеином, богатым аминокислотами с сульфгидрильными и дисульфидными группами. Этот вид нейросекрета наиболее широко распространен у беспозвоночных животных.
Первичный биосинтез нейросекрета в клетках происходит в приядерной зоне цитоплазмы. В перинуклеарной области он появляется в виде мелких пылевидных зернышек, которые затем распространяются по всей клетке (рис. 3). Образование нейросекрета связано с уменьшением размеров ядра и ядрышка, а также заметной редукцией субстанции Ниссля. Электронномикро-скопические исследования показывают, что в синтезе нейросекрета главную роль играет эргастоплазма с ее системой рибосом и аппарат Гольджи (Шаррер и др.). Считают, что образование нейросекрета в клетках ядер гипоталамуса протекает по апокриновому, меракриновому и голокриновому типам (Поленов).
Интенсивность синтеза секрета и выведения его из клеток меняется в зависимости от времени года, условий температурного и светового режимов, физиологического состояния организма, стадии полового цикла и т. д. При дегидратации организма,
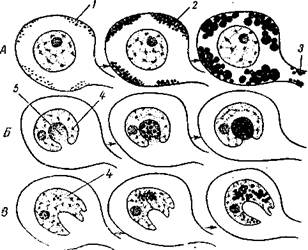 Рис. 74. Последовательные стадии образования нейро-секрета в клетках гипоталамуса
(по Шарреру):
Рис. 74. Последовательные стадии образования нейро-секрета в клетках гипоталамуса
(по Шарреру):
1 — тельца Ниссля; 2 — гранулы секрета; 3 — аксон; 4 — ядро; 5 — базофильная цитоплазма.
например, в клетках паравентрикулярного и супраоптического ядер значительно уменьшается содержание нейросекреторного вещества.
Возрастные изменения гипоталамической нейросекреции изучены еще недостаточно. Образование нейросекрета в супраопти-ческих и паравентрикулярных ядрах гипоталамуса начинается уже в ранний период индивидуального развития животных. У крупного рогатого скота, например, в ядрах подбугорья нейро-секрет обнаруживается у 3-месячных зародышей. У кур первые признаки нейросекреции в супраоптических ядрах обнаруживаются в ранний период инкубации. По данным Денисьевского, образование нейросекрета в клетках паравентрикулярных ядер зародыша уток начинается на 17-й день инкубации. У морских свинок нейросекреторный процесс начинается на 21—28-й день после рождения. У многих животных образование нейросекрета наступает в невронах супраоптического ядра раньше, чем в пара-вентрикулярном. В ранний период развития в гипоталамусе содержится меньше нейросекрета, чем у взрослых животных.
Нейросекрет оказывает физиологическое действие через гуморальную среду организма. В связи с этим значительный интерес представляет вопрос о путях его выведения из нейронов ядер гипоталамуса. Микроскопическими исследованиями установлено, что из невронов супраоптических и паравентрикулярных ядер гранулы гомориположительного нейросекрета по длинным аксонам гипоталамо-гипофизарного тракта токами аксоплазмы смещаются в заднюю долю гипофиза. По мере передвижения от места образования до окончания нервных отростков изменяются химические и тинкториальные свойства нейросекрета. По данным отдельных исследователей, нейросекрет может синтезироваться и в терминалях аксонов, лежащих в задней доле гипофиза (Ди-пен). Задняя доля гипофиза является, таким образом, резервуаром для гипоталамического гомориположительного нейросекрета. Отсюда нейросекрет поступает в кровеносное русло. Часть нейро-секреторного вещества может попадать в кровеносное русло и через ликвор третьего желудочка. Кроме того, нейросекреторные клетки посылают свои аксоны к обонятельным ядрам и эпендиме боковых желудочков переднего мозга. Депонированный в задней доле гипофиза, гомориположительный нейросекрет является носителем физиологически высокоактивных гормонов вазопрессина и окситоцина. По мнению некоторых авторов,_окситоцин образуется в паравентрикулярном, а вазопресин в супраоптическом ядрах. Другие же исследователи считают, что, в зависимости от физиологического состояния организма, клетки одного и того же ядра могут синтезировать оба гормона.
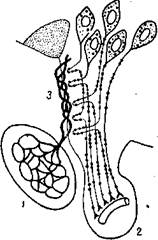 Рис. 75. Портальная система
Рис. 75. Портальная система
сосудов гипофиза у птиц (по
Глису):
1 — передняя доля гипофиза; 2 — задняя доля гипофиза; 3 — срединное возвышение.
Гипоталамический нейросекрет поступает также в переднюю долю гипофиза и контролирует ее гормональную функцию. Однако сюда он поступает из подбугорья не по нервным окончаниям, а с током крови, циркулирующей по сосудам портальной системы. Гуморальный путь тра-нспортировки вырабатываемых клетками гипоталамуса веществ в переднюю долю гипофиза доказывается прямыми экспериментами. У уток, например, портальные вены из гипоталамуса в аденогипофиз проходят обособленно от гипофизарной ножки (рис. 4). Бенуа и Ассенмахер указывают, что перерезка только одной ножки не оказывает существенного влияния на гормональную функцию гипофиза. Перерезка же портальной системы сосудов, при сохранении целостности гипофизной ножки, приводит к угнетению физиологической активности передней доли гипофиза. Показано также, что кровь, взятая из портальных сосудов, стимулирует гипофиз, тогда как кровь из сонной артерии этими свойствами не обладает. Эти эксперименты показывают, что из гипоталамуса в переднюю долю гипофиза действительно поступают вещества, которые активируют его гормональную деятельность. Однако природа этих веществ изучена еще недостаточно.
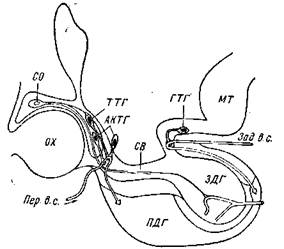
Рис. 5. Схема локализации зон в гипоталамусе собаки, ответственных
за рефляцию тиреотропной (ТТГ), адренокортикотропной (АКТГ) и гонадо-
троной (ГТГ) функции гипофиза (по Хьюму):
Пер. в. с. — передние воротные сосуды; Зад. в. с.—задние воротные сосуды; МТ —
маммилярпос тело; X — хиазма; СО — супраоптическое ядро; ПДГ — передняя доля
гипофиза; ЗДГ — задняя доля гипофиза
Микроскопическими исследованиями установлено, что во внутреннем слое срединного возвышения амиэлиновые волокна ту-бероинфундибулярного пучка, берущие начало от клеток ядер серого бугра, своими окончаниями вступают в связь с расположенными здесь короткими петлями и клубочками первичных капилляров портальной системы (см. рис. 2).
В этих вазоневральных синапсах отростки нервных клеток гипоталамуса отдают в кровь портальной системы секреторное вещество, которое поступает затем в паренхиму передней доли гипофиза. Этот нейросекрет, выделяемый ядрами серого бугра, однако, не окрашивается по Гомори. Отдельные исследователи отождествляли его с медиаторами нервных клеток (ацетилхоли-ном и норадреналином). В настоящее время эта теория уже почти совсем не имеет сторонников. Исследования показали, что нейро-секрсторное вещество от обычных метаболитов нервных клеток отличается более высокой энзиматической устойчивостью и способностью оказывать физиологическое действие на значительном расстоянии от места своего образования в гипоталамусе. Значительный интерес представляет тот факт, что при электролитическом повреждении отдельных участков срединного возвышения в передней доле гипофиза подавляется образование и выведение в кровь не всех, а только какого-либо одного гормона, активирующего функцию строго определенной периферической эндокринной железы (рис. 5). На этом основании считают, что отдельные нервные волокна от различных клеток гипоталамуса несут в срединное возвышение и отдают в кровь портальной системы не одно, а несколько особых веществ или нейрогуморов (Каррато и др.), которые реализуют различные функции передней доли гипофиза (гонадотропную, тиреотропную и адренокор-тикогропную). Возможно, различные нейрогуморы сосудами портальной системы проецируются на определенные зоны паренхимы передней доли гипофиза.
Как уже отмечалось, значение гомориположительного нейро-секрета в регуляции гормональной функции гипофиза изучено еще недостаточно. Обычными методами окрашивания его не удается выявить в туберо-гипофизарном пути и в крови портальной системы. Вместе с тем вокруг капилляров, в их эндотелии и между секреторными клетками передней доли гипофиза часто накапливается значительное количество гранул, которые проявляют такие же реакции, как и гипоталамический неиросекрет. При избыточном введении в организм поваренной соли происходит быстрое выведение нейросекрета из задней доли гипофиза, супраоптического и паравентрикулярного ядер. В этих условиях нейросекретом обогащаются и секреторные клетки передней доли гипофиза (Войткевич и др.). Отдельные исследователи допускают, что в осуществлении связи гипоталамуса с гипофизом важную роль могут играть вазопресин и окситоцин, носителями которых является неиросекрет паравентрикулярного и супраоптического ядер (Мартини и др.). Следует, однако, отметить, что методом хроматографии Саффрон удалось отделить в экстрактах задней доли гипофиза от вазопресина и окситоцина вещество, которое активирует адренокортикотропную функцию передней доли гипофиза.
Приведенные данные показывают, что вопрос о природе нейросекрета, ответственного за регуляцию функции гипофиза, еще нуждается в дальнейшей разработке. Тем не менее, обширный фактический материал указывает на ведущее значение в гнпота-ламическом контроле функции эндокринной системы васкулярных связей. Срединное возвышение серого бугра гипоталамуса и является тем участком, во внутреннем слое которого через посредство вазоневральных синапсов осуществляется передача влияний с гипоталамуса на переднюю долю гипофиза.
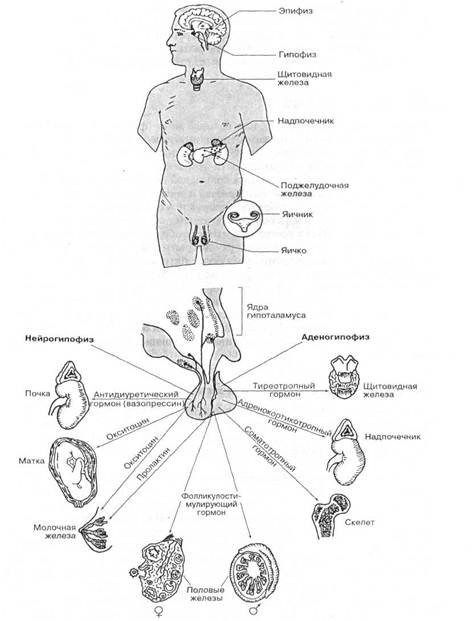
Значение гипоталамуса в регуляции функции эндокринной системы. Гипоталамус принимает участие в нервной и гуморальной регуляции физиологических функций организма. Особенно велико его значение в контроле гормональной деятельности эндокринной системы. Прежде всего гипоталамус сам продуцирует вещества, которые гуморальным путем влияют на отдельные функции организма. Уже отмечалось, что нейроны супраоптического и паравентрикулярного ядер подбугорья синтезируют неиросекрет, который перемещается по нервным отросткам гипоталамо-гипофизарного тракта и аккумулируется в задней доле гипофиза. Этот нейросекрет является носителем физиологически высокоактивных гормонов вазопресина и окситоцина.
Клинические наблюдения и многочисленные экспериментальные исследования последних лет показывают, что гипоталамус оказывает доминирующее влияние на гормональную деятельность передней доли гипофиза и через нее на многие периферические железы внутренней секреции. Этот вывод основан прежде всего на экспериментах по нарушению анатомической связи между гипоталамусом и гипофизом. Так, при перерезке у кроликов (Гаррис), кур (Ширм и Налбандон), уток (Бенуа и Ассен-махер) и других животных гипофизарной ножки резко понижается выделение гипофизом в кровь кринотропных гормонов, активирующих функцию половых желез, коры надпочечников и щитовидной железы. При нарушении связи между гипофизом и гипоталамусом периферические железы переходят в состояние физиологической депрессии. Эта операция особенно сильно отражается на функциональном состоянии половых желез. Если после перерезки ножки портальные сосуды регенерируют и восстанавливается транспортировка из гипоталамуса нейросекрста, то тогда опять нормализуется работа передней доли гипофиза и периферических желез.
Представления о механизмах передачи регулирующих влияний с гипоталамуса на гипофиз за короткую историю разработки этой важной проблемы современной эндокринологии претерпели существенные изменения. На первых этапах ее разработки многие исследователи считали, что влияние гипоталамуса на гипофиз осуществляется через посредство парасимпатической и симпатической нервной системы. Поскольку, однако, прямая нервная связь между ними, по-видимому, отсутствует, то Шаррер уже давно высказал предположение, что гипоталамическая регуляция гормональных функций гипофиза осуществляется преимущественно гуморальным путем при участии нейросекрета. Это положение в дальнейшем подтвердилось не только в опытах с перерезкой портальных сосудов, но и в экспериментах по трансплантации гипофиза в различные органы. При пересадке его гипофизэктомированным животным в почку или в височную долю (Гаррис и Якобсон) гипофиз приживляется, васкуляризируется, но в этих условиях угнетается его гормональная деятельность. Если же этот гипофиз затем подсадить в область срединного возвышения, то после врастания портальных сосудов его гормональная деятельность быстро восстанавливается. Такой же результат получен при совместной инкубации вне организма гипофиза с фрагментами гипоталамуса или при добавлении в культуру экстракта из срединного возвышения подбугорья.
Многочисленные экспериментальные исследования подтверждают, что гипоталамический контроль гормональной функции гипофиза действительно осуществляется через кровь уже рассмотренными прежде нейрогуморами (реализующими факторами). В экспериментальных условиях на функцию передней доли гипофиза может оказывать влияние и суммарный экстракт из нейрогипофиза. На этом основании отдельные исследователи допускают, как это видно на схеме Поленова (таблица I), возможность действия на гормонопоэз передней доли гипофиза и нейросекрета, который поступает в кровеносное русло из нейрогипофиза.
Значительный интерес представляет вопрос о локализации в гипоталамусе участков, ответственных за регуляцию различных гормональных функций гипофиза. При его разработке в настоящее время используются различные приемы. Наиболее широкое применение получил метод точечной электрокаугуляции гипоталамуса, осуществляемой с помощью стереотаксического аппарата, позволяющего производить строго координированные передвижения электродов. Следует, однако, отметить, что и применение стереотаксической техники не устраняет затруднений в решении поставленного вопроса о топографической локализации в гипоталамусе различных зон, регулирующих отдельные функции гипофиза, так как входящие в его состав клеточные компоненты находятся в сложных морфологических и функциональных взаимоотношениях между собой и другими отделами нервной системы. Поэтому повреждение одного участка неизбежно приводит к морфологическим и функциональным нарушениям других компонентов системы. Кроме того, в строении и функциональной дифференцировке отдельных частей гипоталамуса наблюдаются и видовые различия. Вследствие этого полученные различными исследователями данные о значении отдельных участков гипоталамуса в регуляции эндокринных функций организма порою носят противоречивый характер. В настоящее время вполне определенно можно говорить лишь о том, что гипоталамус контролирует гонадотропную, тиреотропную и аденокортико-тропную функции передней доли гипофиза. Для осуществления этих функций, по Бенуа, например, необходима целостность зоны переднего гипоталамуса, расположенной под паравентри-кулярным ядром на границе с преоптическим и туберальным участками.
Рассмотренные выше данные показывают, что гипоталамус и гипофиз в морфологическом и функциональном отношении образуют единую гипоталамо-гипофизарную систему, в которой нервные импульсы переключаются на гуморальные. Значительный интерес, представляет вопрос о механизме работы этого своеобразного пульта управления эндокринными функциями организма. Обширный материал экспериментальных исследований позволяет рассматривать гипоталамус, гипофиз и периферические железы (железы-мишени) как звенья единой системы, функциональная деятельность которой подчиняется принципу обратных связей с самонастройкой на оптимальный для данных условий жизни организма режим работы.
Разработке этих вопросов в свое время много внимания уделял М. М. Завадовский. Давно известно, что избыток в крови гормонов желез-мишеней автоматически приводит к угнетению, а их недостаток — к стимуляции соответствующих тронных функций передней доли гипофиза. Причем угнетение тронной функции наступает в результате повышения концентрации в крови гормона железы-мишени, при некоторых условиях, по-видимому, может осуществляться и непосредственно через гипофиз. Обратный же механизм регуляции, т. е. стимуляция тропной функции гипофиза понижением содержания в крови гормона железы-мишени, осуществляется при обязательном участии гипоталамуса. Изменение уровня гормона в крови является, таким образом, сигналом, который воспринимается клетками соответствующих ядер гипоталамуса.
При описании васкуляризации гипоталамуса уже отмечалось, что особенности строения стенок капилляров и их проницаемости для сложных химических соединений обеспечивают высокую чувствительность нейронов подбугорья к гормонам. Факт непосредственного действия на нейроны гормонов доказывается многочисленными экспериментами подсадки в соответствующие зоны гипоталамуса ткани эндокринных желез или аппликации синтетическими гормонами. Например, имплантация с помощью стереотаксического аппарата кристаллов полового гормона угнетает гонадотропную функцию гипофиза и физиологическую активность половой железы. Сходный результат дает и трансплантация кусочков яичника. Таким образом, через посредство гормонов гипоталамус получает информацию об уровне активности желез-мишеней и посылает в гипофиз сигналы, в ответ на которые последний через продукцию соответствующих тройных гормонов устраняет неблагоприятные для организма отклонения в функции эндокринной системы. Экспериментальные исследования вместе с тем показывают, что в некоторых случаях афферентная сигнализация от желез-мишеней к гипоталамусу может передаваться и нервнопроводниковым путем. Настройка рассмотренной системы обратных связей носит динамический характер и изменяется прежде всего в различные периоды онтогенеза.
В регуляции функций эндокринных желез принимают также участие внегипоталамические центры нервной системы и прежде всего ретикулярная формация. Хотя разработка этого вопроса находится еще в начальной стадии, тем не менее уже теперь имеются многочисленные доказательства ее участия в контроле гормональной активности отдельных эндокринных желез. Эксперименты показывают, что при блокировании фармакологическими средствами, частичном повреждении или раздражении ретикулярной формации электрическим током наступают значительные изменения в уровне гормональной активности отдельных эндокринных желез.
Ретикулярной формации приписывается большое значение в механизме передачи к эндокринным железам разнообразных воздействий на организм, идущих из внешней среды. Характерные изменения в гормональной деятельности надпочечников, щитовидной железы и гонад, наступающие под воздействием не обычных раздражителей в так называемых реакциях «напряжения», или «стресса», многие исследователи также связывают с деятельностью ретикулярной формации.
Пути действия ретикулярной формации на периферические эндокринные железы изучены еще недостаточно. Имеющиеся экспериментальные данные пока не позволяют решить вопрос о том, оказывает ли она только общее активирующее действие на гипоталамус и переключает в него информацию от внешней среды и внутренних органов или же и сама принимает непосредственное участие в регуляции физиологической активности периферических эндокринных желез. Последнее предположение подтверждается отдельными наблюдениями. Известно, что после удаления гипофиза гормональная деятельность отдельных эндокринных желез полностью не прекращается, а сохраняется на уровне так называемой базальной активности, для которой характерна суточная ритмичность. Последняя, по-видимому, контролируется ретикулярной формацией. Рассмотренные факты приводят отдельных исследователей к заключению, что импульсы от ретикулярной формации могут достигать периферических желез без участия гипофиза. Таким образом, возможен и парагипо-физарный путь регуляции эндокринных желез. Ретикулярная формация не только оказывает влияние на гуморальную среду организма, но и сама реагирует на ее изменения. Это указывает на возможность участия ретикулярной формации в рассмотренном выше механизме обратных связей.
Похожие работы
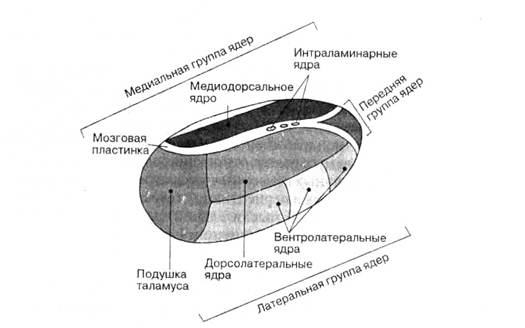
... друг к другу, образуют боковые стенки III мозгового желудочка (полость промежуточного мозга) посередине они соединены между собой межталамическим сращением. Передняя (нижняя) поверхность таламусов сращена с гипоталамусом, через нее с каудальной стороны (т.е. находящейся ближе к нижней части тела) в промежуточный мозг входят проводящие пути из ножек мозга. Латеральная (т.е. боковая) поверхность ...
... гипофиза может иметь различную этиологию (табл. 1463). При наличии несахарного диабета первичный дефект всегда не изменен и локализуется в гипоталамусе или ножке гипофиза, часто сопровождаясь легкой гиперпролактинемией и гипофункцией передней доли гипофиза. Диагноз определяют на основании различных гормональных тестов (см. табл. 1462) и назначают заместительную терапию. Глюкокортикоиды ...

... нервной и эндокринной регуляции в организме // Усп. совр. биол. – 1981. – 19, № 3. – С. 380-382. 23. Вернигора А.Н., Бардинова Ж.С., Сметанин В.А., Генгин М.Т. Активность основных карбоксипептидаз в тканях самок крыс на разных стадиях эстрального цикла // Укр. биохим. журн. – 2003. – 75, № 5. – С. 99-102 24. Вернигора А.Н. Карбоксипептидаза Н мозга животных в норме и при действии стресс- ...
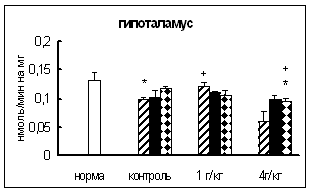
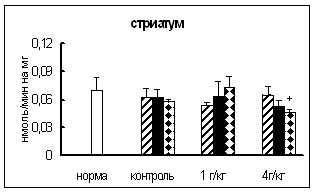
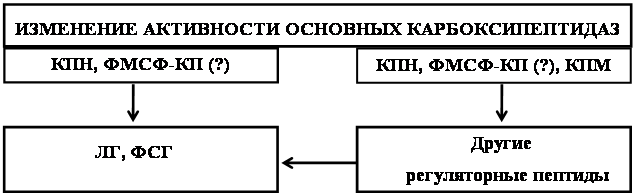
... о динамике изменения активности ферментов. ГЛАВА 3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ 3.1. Исследование активности основных карбоксипептидаз в тканях крыс разного возраста, испытавших пренатальное воздействие этанола 3.1.1. Исследование активности карбоксипептидазы Н в тканях пренатально алкоголизированных крыс разного возраста Согласно данным дисперсионного анализа пренатальное воздействие ...
0 комментариев