Навигация
Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа
31227
знаков
4
таблицы
1
изображение
1. Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа.
Еще в первых опытах на аксоне кальмара было обнаружено, что ионы натрия и калия по-разному влияют на мембранный потенциал. Ионы калия меняют потенциал покоя, а ионы натрия - потенциал действия.
Измерения показали, что ионные каналы обладают абсолютной селективностью по отношению к катионам (катион-селективные каналы), либо к анионам (анион-селективные каналы). В то же время через катион-селективные каналы способны проходить различные катионы различных химических элементов, но проводимость мембраны для неосновного иона, а значит и ток через нее, будет существенно ниже, например, для натриевого канала калиевый ток через него будет в 20 раз меньше. Способность ионного канала пропускать различные ионы называется относительной селективностью и характеризуется рядом селективности - соотношением проводимостей канала для разных ионов, взятых при одной концентрации.
2. Независимость работы отдельных каналов. Прохождение тока через отдельный ионный канал не зависит от того, идет ли ток через другие каналы. Например, калиевые каналы могут быть включены или выключены, но ток через натриевые каналы не меняется. Влияние каналов друг на друга происходит опосредованно: изменение проницаемостей каких-либо каналов (например натриевых) меняет мембранный потенциал, а уже он влияет на проводимости прочих ионных каналов.
3. Дискретный характер проводимости ионных каналов. Ионные каналы представляют собой субъединичный комплекс белков, пронизывающий мембрану. В центре его существует трубка, сквозь которую могут проходить ионы. Количество ионных каналов на 1 мкм поверхности мембраны определяли с помощью радиоактивно-меченного блокатора натриевых каналов - тетродотоксина. Известно, что одна молекула ТТХ связывается только с одним каналом. Тогда измерение радиоактивности образца с известной площадью позволило показать, что на 1 мкм аксона кальмара находится около 500 натриевых каналов. Впервые это было обнаружено в 1962 г. в исследованиях проводимости бислойных липидных мембран (БЛМ) при добавлении в раствор, омывающий мембрану, микроколичеств некоторого вещества, индуцировавшего возбуждение. На БЛМ подавали постоянное напряжение и регистрировали ток. Запись тока во времени имела вид скачков между двумя проводящими состояниями.
Результаты экспериментов выполненных на различных ионных каналах показали, что проводимость ионного канала дискретна и он может находиться в двух состояниях: открытом или закрытом. Выбросы тока обусловлены одновременным открытием 2-х или 3-х каналов. Переходы между состояниями ионного канала происходят в случайные моменты времени и подчиняются статистическим закономерностям. Нельзя сказать, что данный ионный канал откроется именно в этот момент времени. Можно лишь сделать утверждение о вероятности открывания канала в определенном интервале времени.
Ионные каналы описывают характерными временами жизни открытого и закрытого состояний.
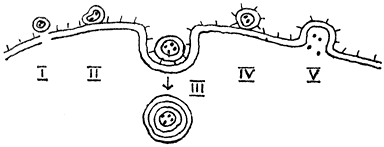
4. Зависимость параметров канала от мембранного потенциала. Ионные каналы нервных волокон чувствительны к мембранному потенциалу, например натриевый и калиевый каналы аксона кальмара. Это проявляется в том, что после начала деполяризации мембраны соответствующие токи начинают изменяться с той или иной кинетикой. На языке "ионных каналов" этот процесс происходит следующим образом. Ион-селективный канал имеет так называемый "сенсор" -некоторый элемент своей конструкции, чувствительный к действию электрического поля (см. рисунок). При изменении мембранного потенциала меняется величина действующей на него силы, в результате эта часть ионного канала перемещается и меняет вероятность открывания или закрывания "ворот" -своеобразных заслонок, действующих по закону "все или ничего".
Структура ионного канала
Ион-селективный канал состоит из следующих частей (см. рисунок): погруженной в бислой белковой части, имеющей субъединичное строение; селективного фильтра, образованного отрицательно заряженными атомами кислорода, которые жестко расположены на определенном расстоянии друг от друга и пропускают ионы только определенного диаметра; воротной части.
"Ворота" ионного канала управляются мембранным потенциалом и могут находиться как в закрытом состоянии (штриховая линия), так и в открытом состоянии (сплошная линия). Нормальное положение ворот натриевого канала -закрытое. Под действием электрического поля увеличивается вероятность открытого состояния, ворота открываются и поток гидратированных ионов получает возможность проходить сквозь селективный фильтр.
Если ион "подходит" по диаметру, то он сбрасывает гидратную оболочку и проскакивает на другую сторону ионного канала. Если же ион слишком велик по диаметру, как например, тетраэтиламмоний, он не в состоянии пролезть сквозь фильтр и не может пересечь мембрану. Если же, напротив, ион слишком мал, то у него возникают сложности в селективном фильтре, на сей раз связанные с трудностью сбросить его гидратную оболочку. У "подходящего" иона сброшенная вода замещается на связи с атомами кислорода, расположенными в фильтре, у "неподходящего" иона стерическое соответствие хуже. Поэтому ему труднее пройти через фильтр и проводимость канала для него ниже.
Блокаторы ионных каналов либо не могут пройти сквозь него, застревая в фильтре, либо, если это большие молекулы как ТТХ, они стерически соответствуют какому-либо входу в канал. Так как блокаторы несут положительный заряд, их заряженная часть втягивается в канал к селективному фильтру как обычный катион, а макромолекула закупоривает его.
Таким образом, изменения электрических свойств возбудимых биомембран осуществляется с помощью ионных каналов. Это белковые макромолекулы, пронизывающие липидный бислой, которые могут находиться в нескольких дискретных состояниях. Свойства каналов, селективных для ионов калия, натрия и кальция могут по-разному зависеть от мембранного потенциала, что и определяет динамику потенциала действия в мембране, а также отличия таких потенциалов в мембранах разных клеток.
* * *
Нельзя преувеличить роль проницаемости плазматической мембраны в жизнедеятельности клетки. Большинство процессов, связанных с обеспечением клетки энергией и избавлением ее от продуктов распада, основаны на вышеописанных механизмах. Кроме того, специальные функции клеточной мембраны заключаются в получении клеткой внешних сигналов (примером этому могут служить описанные взаимодействия клетки с гормонами).
Список использованной литературы:
Антонов В. Ф., Черныш А. М., Пасечник В. И., и др. Биофизика. М., Арктос-Вика-пресс, 1996
Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др. Гистология. М. Медицина, 1999.
Албертс Б., Брэй Д., Льюис Дж. и др. Молекулярная биология клетки. В 3-х томах. Том 1. М., Мир, 1994.
Филлиппович Ю.Б. Основы биохимии. М., Высшая школа, 1985.
Похожие работы
... и многое другое, без чего немыслима сама жизнь. Все человеческое тело – это мир частиц, находящихся в постоянном движении строго по определенным правилам, подчиняющимся физиологии человека. Коллоидные системы организмов обладают рядом биологических свойств, характеризующих то или иное коллоидное состояние: 2.2 Коллоидная система клеток. С точки зрения коллоидно-химической физиологии ...
... температуру, свет, действие различных электрических и магнитных полей, что связано в первую очередь с аморфной структурой самого слабосшитого полиэлектролита. На коллапс геля также могут оказать влияние вещества, соприкасающиеся с сеткой данного геля. Отметим, что его структура может быть глобулярной, которая может разворачиваться при некотором изменении условий. Для изучения факторов, влияющих ...
... информационной плотности, что весьма важно для развития современных технических средств записи, накопления и хранения информации. 7. Важнейшие открытия в химии XXI века 2001 Уильям Ноулз, Риоджи Нойори и Барри Шарплесс «За исследования, используемые в фармацевтической промышленности - создание хиральных катализаторов окислительно-восстановительных реакций». 2002 Джон Фенн и Койчи Танака «За ...
0 комментариев