Навигация
Проприоцептивные пути (от лат. proprius — собственный) проводят импульсы от
149800
знаков
0
таблиц
3
изображения
2. Проприоцептивные пути (от лат. proprius — собственный) проводят импульсы от
органов движения (мышцы, сухожилия, суставные капсулы, связки), несут информацию
о положении частей тела, о размахе движений.
3. Интероцептивные пути (лат. interior — внутренний) проводят импульсы от
внутренних органов, сосудов, где хемо-, баро- и механорецепторы воспринимают
состояние внутренней среды организма, интенсивность обмена веществ, химизм крови
и лимфы, давление в сосудах.
Экстероцептивные проводящие пути. Проводящий путь болевой и температурной
чувствительности — латеральный спинно-таламический путь состоит из трех нейронов. Чувствительным проводящим путям принято давать название с учетом топографии — места начала и конца второго нейрона. Например, у спинно-таламического пути второй нейрон простирается от спинного мозга, где в заднем роге лежит тело клетки, до таламуса, где аксон этого нейрона образует синапс с клеткой третьего нейрона. Рецепторы первого
(чувствительного) нейрона, воспринимающие чувство боли, температуру, располагаются в коже, слизистых оболочках, а нейрит третьего нейрона заканчивается в коре постцентральной извилины, где находится корковый конец анализатора общей чувствительности. Тело первой чувствительной клетки лежит в спинномозговом узле, а центральный отросток в составе заднего корешка направляется в задний рог спинного мозга и заканчивается синапсом на клетках второго нейрона. Аксон второго нейрона, тело которого лежит в заднем роге, направляется на противоположную сторону спинного мозга через его переднюю серую спайку и входит в боковой канатик, где включается в состав латерального спинно-таламического пути. Из спинного мозга пучок поднимается в продолговатый мозг и располагается позади ядра оливы, а в покрышке моста и среднего мозга лежит у наружного края медиальной петли. Заканчивается второй нейрон латерального спинно-таламического пути синапсом на клетках дорсального
латерального ядра таламуса. Здесь лежат тела третьего нейрона, отростки клеток которого проходят через заднюю ножку внутренней капсулы и в составе веерообразно расходящихся пучков волокон, образующих лучистый венец достигают коры полушария большого мозга, его постцентральной извилины. Здесь они заканчиваются синапсами с клетками четвертого слоя (внутренняя зернистая пластинка). Волокна третьего нейрона чувствительного (восходящего) проводящего пути, соединяющего таламус с корой, образуют таламокорковые пучки таламотеменные волокна.
Проводящий путь осязания и давления, передний спинно-таламический путь, несет импульсы от кожи, где лежат рецепторы, воспринимающие чувство давления и осязания, к коре головного мозга, в постцентральную извилину — место расположения коркового конца анализатора общей чувствительности. Тела клеток первого нейрона лежат в спинномозговом узле, а центральные их отростки в составе заднего корешка спинномозговых нервов направляются в задний рог спинного мозга, где заканчиваются синапсами на клетках второго нейрона. Аксоны второго нейрона переходят на противоположную сторону спинного мозга (через переднюю серую спайку), входят в передний канатик и в его составе направляются вверх, к головному мозгу. На своем пути в продолговатом мозге аксоны этого пути присоединяются с латеральной стороны к волокнам медиальной петли и заканчиваются в таламусе, в его дорсальном латеральном ядре, синапсами на клетках третьего нейрона. Волокна третьего нейрона проходят через
внутреннюю капсулу (заднюю ножку) и в составе лучистого венца достигают четвертого слоя коры постцентральной извилины.
Часть волокон проводящего пути осязания и давления идет в составе заднего канатика спинного мозга вместе с аксонами проводящего пути проприоцептивной чувствительности коркового направления.
Необходимо отметить, что не все волокна, несущие импульсы
осязания и давления, переходят на противоположную сторону в спинном мозге. Этот
переход на противоположную сторону частично осуществляется в продолговатом
мозге.
Проприоцептивные проводящие пути. Проводящий путь проприоцептивной
чувствительности коркового направления называется так, поскольку проводит импульсы мышечно-суставного чувства к коре головного мозга, в постцентральную извилину.Чувствительные окончания (рецепторы) первого нейрона располагаются в мышцах, сухожилиях, суставных капсулах, связках. Сигналы о тонусе мышц, натяжении сухожилий, о состоянии опорно-двигательного аппарата в целом (импульсы проприоцептивной чувствительности) позволяют человеку оценить положение частей тела (головы, туловища, конечностей) в пространстве во время движения и проводить
целенаправленные осознанные движения и их коррекцию. Тела первых нейронов лежат
в спинномозговом узле, центральные отростки в составе заднего корешка направляются в задний канатик, минуя задний рог, а затем уходят вверх в продолговатый мозг к тонкому и клиновидному ядрам. Аксоны, несущие проприоцептивные импульсы, входят в задний канатик начиная с нижних сегментов спинного мозга. Каждый следующий пучок аксонов прилежит с латеральной стороны к уже имеющимся. Таким образом, наружные отделы заднего канатика (клиновидный пучок, пучок Бурдаха) заняты аксонами клеток, осуществляющих проприоцептивную иннервацию в верхнегрудных, шейных отделах тела и верхних конечностей. Аксоны, занимающие внутреннюю часть заднего канатика (тонкий пучок, пучок Голля), проводят проприоцептивные импульсы от нижних конечностей и нижней половины туловища. Центральные отростки первого нейрона заканчиваются синапсами на клетках второго нейрона, тела которых лежат в тонком и клиновидных ядрах продолговатого мозга. Аксоны клеток второго нейрона выходят из этих ядер, дугообразно изгибаются вперед и медиально на уровне нижнего угла ромбовидной ямки и в межоливном слое переходят на противоположную сторону, образуя перекрест медиальных петель. Пучок волокон, обращенных в медиальном направлении и переходящих на другую сторону, получил название внутренних дугообразных волокон, fibrae arcuatae internae, которые являются начальным отделом медиальной петли. Волокна медиальной петли в мосту располагаются в задней его части (покрышке), почти на границе с передней частью (между пучками волокон трапециевидного тела). В покрышке среднего мозга пучок волокон медиальной петли занимает место дорсолатеральнее красного ядра, а заканчивается в дорсальном латеральном ядре таламуса синапсами на клетках третьего нейрона. Аксоны клеток третьего нейрона через заднюю ножку внутренней капсулы и в составе лучистого венца достигают постцентральной извилины.
Часть волокон второго нейрона по выходе из тонкого и клиновидного ядер изгибается кнаружи и разделяется на два пучка. Один пучок — задние наружные дугообразные волокна направляются в нижнюю мозжечковую ножку своей стороны и заканчиваются в коре червя мозжечка. Волокна второго пучка — передние наружные дугообразные волокна уходят вперед, переходят на противоположную сторону, огибают с латеральной стороны оливное ядро и также через нижнюю мозжечковую ножку направляются к коре червя мозжечка. Передние и задние наружные дугообразые волокна несут проприоцептивные импульсы к мозжечку.
Проприоцептивный путь коркового направления также перекрещенный. Аксоны второго
нейрона переходят на противоположную сторону не в спинном мозге, а в продолговатом.
Наряду с проприоцептивным проводящим путем, несущим импульсы к коре большого
мозга, следует назвать проприоцептивные передний и задний спинно-мозжечковые пути. По этим проводящим путям мозжечок получает информацию от расположенных ниже чувствительных центров (спинного мозга) о состоянии опорно-двигательного аппарата, участвует в рефлекторной координации движений, обеспечивающих равновесие тела без участия высших отделов головного мозга (коры полушарий большого мозга).
Задний спинно-мозжечковый путь (пучок Флексига) передает проприоцептивные импульсы от мышц, сухожилий, суставов в мозжечок. Тела клеток первого (чувствительного) нейрона находятся в спинномозговом узле, а нейтральные отростки их в составе заднего корешка направляются в задний рог спинного мозга и заканчиваются синапсами на клетках грудного ядра (ядро Кларка), лежащего в медиальной части основания заднего рога. Клетки грудного ядра являются вторым нейроном заднего спинно-мозжечкового пути. Аксоны этих клеток выходят в боковой канатик своей стороны, в его заднюю часть, поднимаются вверх и через нижнюю мозжечковую ножку входят в мозжечок, к клеткам коры червя. Здесь спинно-мозжечковый путь заканчивается.
Можно проследить системы волокон, по которым импульс из коры червя достигает
красного ядра, полушария мозжечка и даже вышележащих отделов мозга — коры
полушарий большого мозга. Из коры червя через пробковидное и шаровидное ядра
импульс через верхнюю мозжечковую ножку направляется к красному ядру
противоположной стороны (мозжечково-покрышечный путь). Кора червя связана
ассоциативными волокнами с корой полушария мозжечка, откуда импульсы поступают в
зубчатое ядро мозжечка. С развитием высших центров чувствительности и произвольных движении в коре полушарий большого мозга возникли также связи мозжечка с корой, осуществляющиеся через таламус. Таким образом, из зубчатого ядра аксоны его клеток через верхнюю мозжечковую ножку выходят в покрышку моста, переходят на противоположную сторону и направляются к таламусу. Переключившись в таламусе на следующий нейрон, импульс следует в кору большого мозга, в постцентральную извилину.
Передний спинно-мозжечковый путь (пучок Говерса) имеет более сложное строение, чем задний, поскольку проходит в боковом канатике противоположной стороны, возвращаясь в конечном итоге в мозжечок на свою сторону. Тело клетки первого нейрона
располагается в спинномозговом узле. Его периферический отросток имеет окончания
(рецепторы) в мышцах, сухожилиях, суставных капсулах, а центральный отросток в
составе заднего корешка входит в спинной мозг и заканчивается синапсами на клетках, примыкающих с латеральной стороны к грудному ядру. Аксоны клеток этого второго нейрона переходят через переднюю серую спайку в боковой канатик противоположной стороны, его переднюю часть, и поднимаются вверх до уровня перешейка ромбовидного мозга. В этом месте волокна переднего спинно-мозжечкового пути возвращаются на свою сторону и через верхнюю мозжечковую ножку вступают в кору червя своей стороны, в его передневерхние отделы. Таким образом, передний спинно-мозжечковый путь, проделав сложный, дважды перекрещенный путь, возвращается на ту же сторону, на которой возникли проприоцептивные импульсы.
Проприоцептивные импульсы, поступившие в кору червя по переднему
спинно-мозжечковому проприоцептивному пути, также передаются в красное ядро и
через зубчатое ядро в кору большого мозга (в постцентральную извилину).
НИСХОДЯЩИЕ ПРОЕКЦИОННЫЕ ПУТИ
Нисходящие проекционные пути (эффекторные, эфферентные) проводят импульсы от
коры, подкорковых центров к нижележащим отделам, к ядрам мозгового ствола и
двигательным ядрам передних рогов спинного мозга. Эти пути можно подразделить на
две группы: 1) главный двигательный, или пирамидный, путь (корково-ядерный и корково-спинномозговые пути) несет импульсы произвольных движений из коры головного мозга к скелетным мышцам головы, шеи, туловища, конечностей через соответствующие двигательные ядра головного и спинного мозга;
2) экстрапирамидные двигательные пути передают импульсы от подкорковых центров к двигательным ядрам черепных и спинномозговых нервов, а затем к мышцам.
К пирамидному пути относится система волокон, по которым двигательные импульсы из коры большого мозга, из предцентральной извилины, от гигантопирамидальных нейронов (клетки Беца) направляются к двигательным ядрам черепных нервов и передним рогам спинного мозга а от них — к скелетным мышцам. Учитывая направление хода волокон, а также расположение пучков в стволе головного мозга и канатиках спинного мозга, пирамидный путь подразделяют на три части: 1) корково-ядерный — к ядрам черепных нервов; 2) латеральный корково-спинномозговой (пирамидный) — к ядрам передних рогов спинного мозга; 3) передний корково-спинномозговой (пирамидный) — также к
передним рогам спинного мозга.
Корково-ядерный путь представляет собой пучок отростков гигантопирамидальных нейронов, которые из коры нижней трети предцентральной извилины спускаются к внутренней капсуле и проходят через ее колено. Далее волокна корково-ядерного пути идут в основании ножки мозга, образуя медиальную часть пирамидных путей. Корково-спинномозговые, а также корково-ядерный пути занимают средние 3/5 основания ножки мозга. Начиная со среднего мозга и далее, в мосту и продолговатом мозге волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов III и IV — в среднем мозге, V, VI, VII — в мосту, IX, X, XI, XII — в
продолговатом мозге. В этих ядрах корково-ядерный (пирамидный) путь заканчивается, составляющие его волокна образуют синапсы с двигательными клетками этих ядер. Отростки упомянутых двигательных клеток выходят из мозга в составе соответствующих черепных нервов и направляются к скелетным мышцам головы и шеи и их иннервируют.
Латеральный и передний корково-спинномозговые (пирамидные) пути, также начинаются от гигантопирамидальных нейронов предцентральной извилины, ее верхних 2/3. Аксоны этих клеток направляются к внутренней капсуле, проходят через переднюю часть ее задней ножки (сразу позади волокон корково-ядерного пути), спускаются в основание ножки мозга, где занимают место латеральнее корково-ядерного пути. Далее корково-спинномозговые волокна спускаются в переднюю часть (основание) моста, пронизывают идущие в поперечном направлении пучки волокон моста и выходят в продолговатый мозг, где на передней (нижней) его поверхности образуют выступающие вперед валики — пирамиды. В нижней части продолговатого мозга часть волокон переходит на противоположную сторону и продолжается в боковой канатик спинного мозга, постепенно заканчиваясь в передних рогах спинного мозга синапсами на двигательных клетках его ядер. Эта часть пирамидных путей, участвующая в образовании перекреста пирамид (моторный перекрест), получила название латерального корково-спинномозгового (пирамидного) пути. Те волокна корково-спинномозгового пути, которые не участвуют в образовании перекреста пирамид и не переходят на противоположную сторону, продолжают свой путь вниз в составе переднего канатика спинного мозга. Эти волокна составляют передний корково-спинномозговой (пирамидный) путь. Затем эти волокна также переходят на противоположную сторону, но через белую спайку спинного мозга и заканчиваются на двигательных клетках переднего рога противоположной стороны спинного мозга. Располагающийся в переднем канатике
передний корково-спинномозговой (пирамидный) путь более молодой в эволюционном
плане, чем латеральный. Его волокна спускаются преимущественно до уровня шейных
и грудных сегментов спинного мозга. Следует отметить, что все пирамидные пути
являются перекрещенными, т.е. их волокна на пути к следующему нейрону рано или
поздно переходят на противоположную сторону
Вторым нейроном нисходящего произвольного двигательного пути (корково-спинномозгового) являются клетки передних рогов спинного мозга, длинные отростки которых выходят из спинного мозга в составе передних корешков и направляются в составе спинномозговых нервов для иннервации скелетных мышц.
Экстрапирамидные проводящие пути, объединенные в одну группу, в отличие от более
новых пирамидных путей, являются филогенетически более старыми, имеющими
обширные связи в мозговом стволе и с корой большого мозга, взявшей на себя функции контроля и управления экстрапирамидной системой. Кора большого мозга, получающая импульсы как по прямым (коркового направления) восходящим чувствительным путям, так и из подкорковых центров, управляет двигательными функциями организма через экстрапирамидные и пирамидные пути. Кора большого мозга оказывает влияние на двигательные функции спинного мозга через систему мозжечок-красные ядра, через ретикулярную формацию, имеющую связи с таламусом и полосатым телом, через вестибулярные ядра. Таким образом, в число центров экстрапирамидной системы входят красные ядра, одной из функций которых является поддержание мышечного тонуса, необходимого для удерживания тела в состоянии равновесия без усилия воли. Красное ядро, которое относится также к ретикулярной формации, получает импульсы из коры большого мозга, мозжечка (от мозжечковых проприоцептивных путей) и само имеет связи с двигательными ядрами передних рогов спинного мозга.
Красноядерно-спинномозговой путь входит в состав рефлекторной дуги, приносящим звеном которой являются спинно-мозжечковые проприоцептивные проводящие пути. Этот путь берет начало от красного ядра (пучок Монакова), переходит на противоположную сторону (перекрест Фореля) и спускается в боковом канатике спинного мозга, заканчиваясь на двигательных клетках спинного мозга. Волокна этого пути проходят в задней части (покрышка) моста и боковых отделах продолговатого мозга.
Важным звеном в координации двигательных функций тела человека является
преддверно-спинномозговой путь. Он связывает ядра вестибулярного аппарата с передними рогами спинного мозга и обеспечивает установочные реакции тела при нарушении равновесия. В образовании преддверно-спинномозгового пути принимают участие аксоны клеток латерального вестибулярного ядра (ядро Дейтерса), а также нижнего вестибулярного ядра (нисходящего корешка) преддверно-улиткового нерва. Эти волокна спускаются в латеральной части переднего канатика спинного мозга (на границе с боковым) и заканчиваются на двигательных клетках передних рогов спинного мозга. Ядра, образующие преддверно-спинномозговой путь, находятся в непосредственной связи с мозжечком, а также с задним продольным пучком, который в свою очередь связан с ядрами глазодвигательных нервов. Наличие связи с ядрами глазодвигательных нервов обеспечивает сохранение положения глазного яблока (направление зрительной оси) при поворотах головы и шеи. В образовании заднего продольного пучка и тех волокон, которые достигают передних рогов спинного мозга (ретикулярно-спинномозговой путь) принимают участие клеточные скопления ретикулярной формации стволовой части мозга, главным образом промежуточное ядро, (ядро Кахаля), ядро эпиталамической (задней)
спайки (ядро Даркшевича), к которым приходят волокна из базальных ядер полушарий большого мозга.
Управление функциями мозжечка, участвующего в координации движений головы, туловища и конечностей и связанного в свою очередь с красными ядрами и
вестибулярным аппаратом, осуществляется из коры большого мозга через мост по
корково-мостомозжечковому пути. Этот проводящий путь состоит из двух нейронов. Тела клеток первого нейрона лежат в коре лобной, височной, теменной и затылочной долей. Их отростки — корково-мостовые волокна направляются к внутренней капсуле и проходят через нее. Волокна из лобной доли, которые можно назвать лобномостовыми волокнами, проходят через переднюю ножку внутренней капсулы, нервные волокна из височной, теменной и затылочной долей — через заднюю ножку. Далее волокна корково-мостовых путей идут через основание ножки мозга. От лобной доли волокна проходят через самую медиальную часть основания, кнутри от корково-ядерных волокон, а от теменной и других долей — через самую латеральную часть, кнаружи от корково-спинномозговых путей. В передней части (в основании) моста волокна корково-мостовых путей заканчиваются синапсами на клетках ядер моста этой же стороны мозга. Клетки ядер моста с их отростками составляют второй нейрон корково-мостомозжечкового пути. Аксоны клеток ядер моста складываются в пучки — поперечные волокна моста, которые переходят на противоположную сторону, пересекают при этом в поперечном направлении нисходящие пучки волокон пирамидных путей и через среднюю мозжечковую ножку направляются в полушарие мозжечка противоположной стороны.
Таким образом, проводящие пути головного и спинного мозга устанавливают связи
между афферентными и эфферентными (эффекторными) центрами, участвуют в образовании сложных рефлекторных дуг в теле человека. Одни проводящие пути
(системы волокон) начинаются или заканчиваются в филогенетически более старых,
лежащих в мозговом стволе ядрах, обеспечивающих функции, обладающие определенным
автоматизмом. Эти функции (например, тонус мышц, автоматические рефлекторные
движения) осуществляются без участия сознания, хотя и под контролем коры большого мозга. Другие проводящие пути передают импульсы в кору большого мозга, в высшие отделы ЦНС или из коры к подкорковым центрам (к базальным ядрам, ядрам мозгового ствола и спинного мозга). Проводящие пути функционально объединяют организм в одно целое, обеспечивают согласованность его действий.
Список литературы:
1.Сапин М.Р. « Анатомия» М: «Высшая школа» ,1989 г.
2.Михеев в.В. « Нервные болезни» М: «Медицина» ,1973 г.
3. Синельников Р.Д. «Атлас анатомии человека» т. 1,3 М: «Медицина» ,1968 г.
4. «Малая медицинская энциклопедия» т.6 М: «Советская энциклопедия» ,1967 г.
Похожие работы
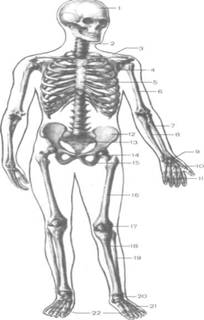
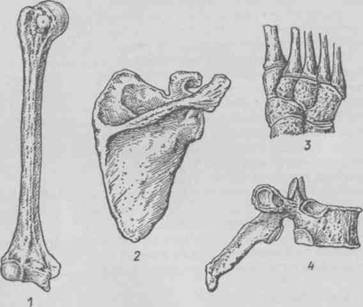
... суставных капсул, связок. Формирование суставных поверхностей, капсул и связок завершается в основном в подростковом возрасте (13—16 лет). 4. МЫШЕЧНАЯ СИСТЕМА 4.1 Строение мышцы Скелетные мышцы являются активной частью опорно-двигательного аппарата, построены они из поперечнополосатых (исчерченных) мышечных волокон. Мышцы прикрепляются к костям скелета и при своем сокращении ( ...
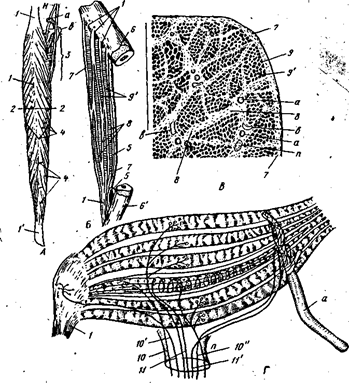

... увеличиваются диаметры мышечных волокон и их количество. Только двигательная активность при откорме приводит к увеличению массы тела животного за счет мышечной, а не соединительной ткани или жира. Не надо забывать, что мышцы во время динамической нагрузки работают еще и как «периферическое сердце», обеспечивая норму кровообращения, а стало быть, и норму обмена веществ в организме. Характеристика ...
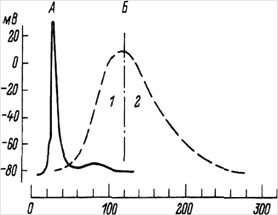
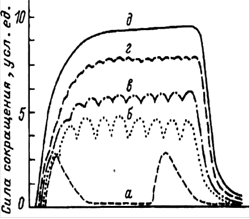
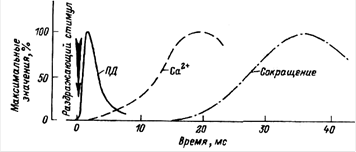
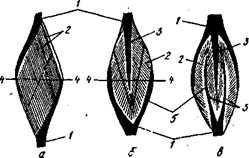
... ) наносится либо во время 2-й фазы (расслабления или удлинения), либо во время 1-й фазы (укорочения или напряжения). Одиночное сокращение А - потенциал действия; Б - сокращение мышцы 1 - фаза напряжения; 2 - фаза расслабления Суммированное сокращение а - одиночное сокращение; б-г - зубчатый тетанус; д - гладкий тетанус В случае, когда всякое второе раздражение ...
... В.С. Фарфель и др. Нервная система едина, но условно ее делят на части. Имеется две классификации: по топографическому принципу, т. е. по месту расположения нервной системы в организме человека, и по функциональному принципу, т. е. по областям ее иннервации. По топографическому принципу нервную систему делят на центральную и периферическую. К центральной нервной системе относят головной мозг и ...
0 комментариев