Навигация
Поведение в вероятностной среде
37197
знаков
0
таблиц
0
изображений
3. Поведение в вероятностной среде.
Понимание того, что животное и человек, как правило, действуют в постоянно и случайно меняющейся среде, побудило исследователей к изучению способности нервной системы отражать вероятные характеристики действительности.
Способность живого организма прогнозировать вероятность предстоящих событий показана во многих экспериментах на крысах, кошках, обезьянах и человеке. При этом исследователи подчеркивают особую роль передних отделов новой коры. Так, после повреждения лобных долей у обезьян нарушение отражения вероятных характеристик среды сохраняется в течение трех с лишним лет, тогда как аналогичный дефект, возникающий после двухстороннего удаления теменной коры, исчезает через один – полтора месяца.
Вместе с тем способность к отражению вероятной структуры среды показана и для других структур мозга. В опытах Д.Г.Шевченко в лаборатории В.Б.Швыркова это свойство было установлено у нейронов зрительной коры. Кролика обучали в ответ на вспышки света слева и справа нажимать на педаль и получать пищу соответственно из левой и правой кормушки. Вспышки света с разных сторон подавались с разной вероятностью: 50% и 100%. Нейроны зрительной коры научились различать вероятность проявления вспышек, что выразилось в формировании у них различных реакций. Эти дифференцированные реакции нейронов появлялись перед нажимом на педаль. И не выявлялись на других этапах поведения.
Нервная система обладает способностью оценивать не только вероятность появления тех или других стимулов, но и вероятность удовлетворения потребности, т. е. вероятность с которой следует подкрепление. П.В.Симонов различает два самостоятельных механизма, измеряющих вероятность подкрепления, связывая их с функциями различных структур мозга. По его мнению, реагирование на условные сигналы, подкрепляемые с низкой вероятностью, обеспечивается гиппокампом, тогда как прогнозирование высоковероятностного подкрепления связано с функциями фронтальной коры.
Это заключение строиться на результатах опытов, в которых показано, что после разрушения передних отделов новой коры избирательно замедляется процесс формирования условных пищевых и двигательных реакций только при высокой вероятности подкрепления. Выработка условных рефлексов при низкой вероятности подкрепления не страдает, в результате чего значение редко и часто подкрепляемых условных стимулов выравнивается. Повреждение теменной коры у кошки не нарушает высоковероятностное прогнозирование.
Исследователи высказывают предположение, что повышенная отвлекаемость на малозначимые события, которая наблюдается у больных с опухолью в лобных долях мозга, может быть связана с нарушением механизмов вероятностного прогнозирования.
В отличие от фронтальной коры гиппокамп необходим для реагирования на сигналы с низкой вероятностью подкрепления. По данным М.Я.Пигаревой, двустороннее удаление гиппокампа у крыс делает невозможным их обучение при подкреплении с вероятностью 0,4 и 0,33. Нейроны гиппокампа фиксируют все изменения, все нестандартные отклонения в окружающей среде, вызывающие ориентировочные реакции. Чем ниже вероятность события, тем сильнее реакция нейронов гиппокампа.
Существуют индивидуальные различия в способности людей прогнозировать вероятность событий. Одни лица имеют тенденцию к субъективному завышению частоты более частого события, другие занижают её, а третьи адекватно оценивают вероятность как частых, так и редких явлений. По некоторым данным, способность отражать вероятностную структуру среды связана преимущественно с функцией правого полушария.
Таким образом, многочисленные данные показывают, что отражение вероятностной структуры среды является одним из важных механизмов работы мозга. Функции этого механизма распределены между различными структурами мозга. Вероятностные характеристики действительности на ряду с другими ее параметрами выделяются нервной системой и фиксируются в памяти животного и человека.
Это обстоятельство дает основание исследователям полагать, что механизм предвидения в структуре поведенческого акта должен строиться с учетом как образа «вероятностной структуры среды», так и «вероятности достижения цели».
Следует отметить, что сам П.К.Анохин возражал против термина «вероятностное прогнозирование» на том основании, что вероятность прогноза, который представлен в акцепторе результатов действия, всегда максимальна и равна единице. В каждый данный момент времени цель, которая уже сформирована в акцепторе действия, всегда одна. Вместе с тем его модель поведения не отрицает способности нервной системы к отражению вероятностных свойств действительности. Согласно вышерассмотренной модели поведенческого акта эта информация вместе с другой на стадии аффентного синтеза извлекается из памяти и используется для формирования акцептора действия. Т.е. отражение вероятностной структуры среды связано с ранними стадиями развертывания поведенческого акта, предшествующими стадии акцептора результатов действия.
По мнению А.С.Батуева, знания вероятностных характеристик среды, которому он предает очень важное значение в построении поведения, определяет степень актуальности той или другой двигательной программы, извлекаемой из памяти. Отдельные поведенческие программы выстраиваются в последовательность в соответствии с вероятностными характеристиками действительности.
Важная функция в структуре поведенческого акта принадлежит ориентировочно – исследовательской деятельности, которая всякий раз возобновляется в результате рассогласования акцептора действия с параметрами и результатом выполненного действия. Именно благодаря росту неспецифической активации в ЦНС, которая наблюдается во время ориентировочных реакций, становятся возможными более тщательный анализ окружающей среды и более успешное извлечение информации из памяти для последующего формирования нового акцептора действия. Значение для поведения механизма, регулирующего уровень активации нервной системы, выходит далеко за пределы обеспечения ориентировочно – исследовательской деятельности, так как сама реализация поведения в соответствии с содержанием акцептора действия сильно зависит от процессов неспецифической активации.
4.Нейронные механизмы поведения.
Переход исследователей к изучению нейронных механизмов поведения открывает широкие перспективы как для понимания назначения отдельных структур мозга, так и самой организацией поведенческого акта. О чем же говорит изучение нейронной активности?
Любой поведенческий акт представляет интеграцию врожденных и приобретенных составляющих, от соотношения которых зависит степень его сложности. Поведенческий же акт, который в основном предопределен наследственностью, генетической памятью, имеет более простую структуру.
На нейронном уровне он может быть представлен как интеграция сенсорных и командных нейронов, которые реализуют моторный акт через пул мотонейронов. Влияния мотивационного и неспецифического возбуждения на такую систему реализуются через модулирующие нейроны.
Поведенческий акт, который формируется в процессе индивидуальной жизни и является результатом обучения, имеет более сложную организацию. Изучение активности отдельных нейронов во время выполнения сложного поведения позволяет выделить большее число групп нейронов, различающихся своими функциями.
Прежде всего выделена большая группа сенсорных нейронов. Среди них есть нейроны – детекторы. Они селективно реагируют на определенное и достаточно простое качество или свойство внешнего мира. Это детекторы элементов формы, выделяющие углы, отрезки линий, определенным образом ориентированные, или детекторы цвета и т.д. Среди сенсорных нейронов выделена группа специальных нейронов – «гностических единиц», избирательно реагирующих уже не на простые, а на сложные интегральные признаки. Так, Е.Роллсом в верхней височной коре у обезьян найдены нейроны, избирательно реагирующие на определенные лица людей или других обезьян, а также на фотографии и не реагирующие на неодушевленные объекты и их изображения. Кроме того, также найдены нейроны, идентифицирующие эмоции, например, они избирательно отвечают на выражение угрозы на человеческом лице. Нейроны, идентифицирующие лица и эмоциональную экспрессию, независимо от ракурса восприятия, обнаружены также в миндалине. Полагают, что данные гностические единицы в миндалине принимают участие в формировании коммуникативного поведения животных в стаде.
Таким образом, реакция организма на пусковую и обстановочную афферентацию предполагает возбуждение различных сенсорных нейронов, включая детекторы простых и сложных признаков.
Особую группу нейронов составляют нейроны «среды» которые избирательно возбуждаются на определённую обстановку, место в привычном сенсорном окружении. К ним относятся пространственно-селективные нейроны, описанные А.С.Батуевым. Эти нейроны предпочитали реагировать на условный сигнал, предъявляемый либо слева, либо справа. Нейроны «среды» были найдены Ю.Н.Сущенко в моторной, соматосенсорной и зрительной коре у кролика. Их активация наблюдалась только в момент пребывания кролика в определенной части клетки. При этом их активность не зависела от каких – либо движений и поведения кролика. Нейроны «среды» в коре сходны с нейронами «места», найденными О`Кифом в гиппокампе кролика. Нейроны «места» также активировались лишь при определенной ориентации животного в экспериментальном пространстве. Некоторые из них имели не одно, а два и более предпочитаемых пространственных полей в окружающей среде животного.
Во многих структурах мозга обнаружены нейроны, активация которых связана с выделением поведенческого акта. Прежде всего это нейроны, которые избирательно реагируют на внешний вид пищи. Они найдены в гипоталамусе, в височной коре обезьян, в хвостатом ядре. А.С.Батуев нашел их в теменной и лобной коре у обезьян. Реакции этих нейронов возникали без обучения. Но их появление зависело от мотивационного возбуждения животного (голода). С насыщением обезьяны эти нейроны переставали реагировать на вид пищи. Одновременно уменьшается число правильных реакций выбора. С заменой подкрепления на более привлекательное реакция активации нейронов на вид пищи восстанавливается, а с ней и общий уровень условнорефлекторной деятельности.
Сходный тир нейронов был обнаружен К.В.Судаковым, который назвал их нейронами «ожидания». В условиях пищевого мотивационного возбуждения, возникающего естественным путем, или в результате электрического раздражения «центра голода», расположенного в латеральном гипоталамусе, эти нейроны разряжаются пачками спайков. С удовлетворением голода пачечный тип активности нейронов замещается на следующие равномерно одиночные спайки.
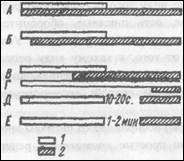
Пачечный тип активности нейрона, характерный для голодного кролика (1), после насыщения животного сменяется частыми, равномерно следующими одиночными спайками (2). Сверху вниз: отметка кормления животного, запись нейронной активности и межимпульсные гистограммы – по оси абсцисс – мс, по оси ординат – число спайков (по К.В.Судакову, 1986).
При попадании пищи в полость рта пачечная активность нейронов усиливается и исчезает в момент поступления пищи в желудок. Следовательно, контактное восприятие пищи через вкусовые рецепторы, т.е. предмета способного удовлетворить голодную мотивацию, что отражается в исчезновении пачечной активности у этих нейронов.
Исследователи высказывают гипотезу, что нейроны, на которых конвергируют мотивационное возбуждение и возбуждение от подкрепления, входят в структуру акцептора результатов действия. Пачечная активность этих нейронов отражает «ожидание» пищевого подкрепления. В нейронах ожидания во время пищевой мотивации закодирована информация о предмете, способном удовлетворить голод, т. е. информация о его виде, вкусе, и способности утолять голод. Мотивационное возбуждение актуализирует образ предмета и тем самым повышает реактивность нейрона на пищевые раздражители. В результате вид пищи вызывает мощную реакцию нейрона в виде усиления пачечной активности. В отсутствие мотивации нейрон не реагирует на вид пищи. Изменение реакций нейронов «ожидания» объясняют, почему голодным животным пища воспринимается иначе, чем вытым. Таким образом, в условиях мотивационного возбуждения возникает пристрастное восприятие предметов, способных удовлетворять потребности. Нейроны «ожидания» широко представлены в коре и подкорке. Число их увеличивается по направлению от коры к стволовым структурам мозга: от 30% в соматосенсорной коре до 75 – 80% в ретикулярной формации среднего и продолговатого мозга. Полагают, что нейроны разных отделов мозга «ожидают» свои специфические параметры подкрепления. Каждой мотивации: пищевой, питьевой, оборонительной и т.п. – соответствует свой тип пачечной активности, свой тип распределения межимпульсных интервалов, одновременно охватывающий многие нейроны самых разных структур мозга.
Выделена также группа нейронов, которые активируются при выполнении целевых движений. В.Б.Швырков назвал их нейронами «цели». Их активация предшествует акту хватания пищи у кроликов либо нажиму на педаль, за которым следует подача кормушки с пищей. Активация этих нейронов наблюдается при любых вариантах приближения к цели (справа, слева) и при любом способе нажатия на педаль (одной или двумя лапами), и она всегда прекращается при достижении результата. Условия, среда, в которой выполняется данное действие, также не влияют на активность нейронов «цели». Нейроны цели зарегистрированы в моторной, сенсомоторной, зрительной коре, гиппокампе кролика. Они не активируются перед пережевыванием пищи, не направленными на достижение цели. Нейроны, которые возбуждались только перед выполнением заученного движения нажима на определенный рычаг, сопровождающегося пищевым подкреплением, были обнаружены и у обезьян в лобной и теменной коре. А.С.Батуев их называет нейронами моторных программ, так как их активация связана с запуском двигательного акта. Активации нейронов моторных программ предшествует возбуждение двух других групп нейронов: 1) в момент действия условного сигнала и 2) в период отсрочки между условным сигналом и моментом, когда обезьяна получает доступ к рычагу.
Наконец, многими исследователями выделяются нейроны, активность которых жестко связана с осуществлением определенных движений независимо от их роли и места в структуре поведенческого акта. Так описаны нейроны, которые активируются при занятии животных определенной позы, при пережевывании пищи. Среди них следует различать командные нейроны и мотонейроны. Возбуждение командного нейрона определяет осуществление сложно координированного движения, выполнение которого вовлекается группа мотонейронов. Возбуждение же мотонейрона связано с сокращением или расслаблением отдельной мышцы.
В результате несоответствия модели ожидаемого будущего с наличным возбуждением на смену основного поведения, направленного на удовлетворение доминирующей потребности, приходит ориентировочно – исследовательское поведение. Так, не обнаружив педали, нажим на которую сопровождается подачей пищи в соответствующую кормушку, кролик активно обследует клетку, обнюхивает новую кормушку и педаль, скребет место старой кормушки лапами. По данным В.Б.Швыркова, ориентировочно – исследовательское поведение в некоторых случаях сменяется оборонительным, агрессивным поведением. При этом кролики зубами вырывали педали и швыряли их по клетке. Такое поведение имеет приспособительное значение: животное использует весь свой индивидуальный опыт независимо от условий его приобретения. Само ориентировочно – исследовательское поведение направлено на детальное ознакомление с окружающей средой и поиск тех сигналов или объектов, которые ранее как – то были связаны с удовлетворением доминирующей потребности. В.Б.Швырковым выделена особая группа нейронов которые становятся активными только в поисковом поведении кролика. Такие нейроны были обнаружены в сенсомоторной коре и головке хвостатого ядра. В его опытах голодный кролик (24ч депривации) мог поедать листья, укрепленные на полу камеры специальных держателях. Нейроны поискового поведения активировались при обнюхивании пустых держателей, но только тех из них, из которых кролик ранее хотя бы один раз уже получал пищу. Эта реакция нейронов во время обнюхивания вырабатывается и может быть угашена. Для этого нужно, чтобы у кролика сменился держатель, через который его подкармливают.
Характерной особенностью ориентировочно – исследовательского поведения является также усиление ориентировочных реакций, обеспечивающих лучшее восприятие стимулов. На нейронном уровне этому соответствует усиление реакций особого класса нейронов – нейронов новизны, активирующихся при действии новых стимулов и снижающих свою активность по мере привыкания к ним. Нейроны «новизны» описаны для гиппокампа, неспецифического таламуса, ретикулярной формации среднего мозга и других структур.
Таким образом, исследования, выполненные на нейронном уровне, приводят к заключению, что активность нейронов связана с поведенческим актом и что отдельные его стадии, этапы представлены различными группами нейронов. При этом функционально сходные нейроны могут быть обнаружены в различных структурах мозга, т.е. существует определенная распределенность функции по различным структурам мозга, что не отрицает их специализации. Так, для гиппокампа характерны нейроны «места», хотя они найдены также в неокортексе, в гипоталамусе. Но в этих структурах они представлены в малых количествах, тогда как в полях гиппокампа они составляют большинство.
Структура поведенческого акта. Нейронные механизмы поведения.
Содержание.
1.Функциональные системы:
а)концепция функциональной системы П.К.Анохина;
б) «функциональная система – единица интегративной деятельности всего организма»;
2.Стадии поведенческого акта:
а)стадия афферентного синтеза;
б)принятия решений;
в)акцептор результата действия;
г)эфферентный синтез;
д)формирование действий и оценка результата.
3.Поведение в вероятностной среде.
а)вероятностное прогнозирование;
б)отражение вероятностной структуры среды;
в)ориентировочно – исследовательская деятельность.
4.Нейронные механизмы поведения.
Список используемой литературы.
1.Н.Н.Данилова, А.Л.Крылова «Физиология высшей нервной деятельности» «Феникс» 1999г.
2.Е.Н.Соколов «Физиология высшей нервной деятельности»М., 1981г.
3.К.В.Судаков «Физиология поведения». Л.,1907г.
4.Ю.И.Александров «Основы психофизиологии»М., 1998г.
Похожие работы




... подход к оценке механизмов работы высших отделов головного мозга с использованием условных рефлексов был столь успешным, что позволил Павлову создать новый раздел физиологии — «Физиологию высшей нервной деятельности», науку о механизмах работы больших полушарий головного мозга. БЕЗУСЛОВНЫЕ И УСЛОВНЫЕ РЕФЛЕКСЫ Поведение животных и человека представляет собой сложную систему взаимосвязанных ...
... афферентных систем представляют собой только часть тех сложных физиологических структур, которые воспринимают различные раздражения, преобразуют их в нервные импульсы, проводят в соответствующие центры ЦНС, где обеспечивается анализ информации. И.П. Павлов закономерно объединил в понятие психической деятельности два механизма: 1 – механизм условных рефлексов, 2 – механизм анализаторов (высших ...
... только один половой инстинкт. Во-вторых, потому, что в человеческом поведении исключительная роль принадлежит социальным реакциям — поведению, определяемому положением человека в обществе, коллективе, социальной среде. К сожалению, зарубежные физиологи зачастую строят свои нейрефизиологические концепции, исходя из психоанализа Фрейда. [функцию коры, общую для человека и животных, обуславливаю- ...
... и утрачивается к началу полового созревания. 36. Учение Павлова о типах ВНД. Общин типы ВНД. Пластичность ВНД. Учет типов ВНД в индивидуальном обучении и воспитании Среди вопросов физиологии ВНД человека особое значение для совершенствования учебно-воспитательного процесса и разработки естественнонаучных теорий воспитания и обучения имеет учение о типах ВНД. Это связано с тем, что именно ...
0 комментариев