как разновидность тканевого диспротеиноза широко распространен среди представителей животного мира. Изучение амилоидоза насчитывает более 100 лет. Однако патогенез амилоидоза сложен и, несмотря на тщательное изучение в последние годы, не все его звенья достаточно изучены на сегодняшний день. Существует определение амилоидоза как такового: амилоидоз - это мезенхимальный диспротеиноз, сопровождающийся нарушением белкового обмена, появлением аномального фибриллярного белка и образованием в межуточной ткани сложного вещества - амилоида.
Амилоид, его природа и свойства.
В 1853 году Вирхов показал, что вещества, выпадающие в тканях при "сальной болезни" Рокитанского, обладают свойством окрашиваться йодом подобно крахмалу и назвал его амилоидом.
Химический состав и физические свойства амилоида.
Амилоид является сложным веществом - гликопротеидом, в котором фибриллярные и глобулярные белки связаны с полисахаридами.
Белки амилоида.
По Кравкову (1894 год) в состав амилоида входят:
углерод (48,86 - 50,38 %);
водород (6,45 - 7,09 %);
азот (13,79 - 14,67 %);
сера (2,65 - 2,8 %);
фосфор (следы).
Гилес и Калкинс (1955 год ) обнаружили в амилоиде 82,8 % воды, до 14,2% азота, до 4% углеводов, 0,9% фосфора и 0,86% серы.
Амилоид дает характерные для белков биуретовую и ксантопротеиновую реакции. Большой процент содержания воды в амилоиде говорит о высокой гидрофильности белков, а низкое содержание фосфора предполагает, что он принадлежит к нуклеиновым кислотам, имеющимся в амилоиде хотя достоверных данных нет). Амилоид пиронинофилен.
Аминокислотный состав амилоида отличается от сывороточных и тканевых белков ( в моль-процентах):
глицин - 10-17,3;
аланин - 9,7-14,4;
лейцин - 7,7-13,7;
валин - 4,1 - 7,1;
метионин - 1,1;
цистеин - 1,5 - 2,9;
тирозин - 2,7-3,7;
триптофан;
гистидин.
В амилоиде преобладают липофильные группы (глицин, аланин, лейцин, валин).
Белковый компонент амилоида сходен с глобулинами. Величины содержания метионина и цистеина оказались близкими к тем, которые были получены при анализе гамма-глобулина сыворотки, хотя содержание аминокислот в амилоиде и в сывороточном гамма-глобулине различен.
Наличие триптофана свидетельствует о его близости к фибриногену, который также содержит эту аминокислоту в большом количестве. Амилоид содержит много тирозина (гиалин и коллаген). Качественно аминокислотный состав амилоида одинаков при различных формах амилоидоза, однако количественная характеристика компонентов, входящих в его состав колеблется в зависимости не только от формы амилоидоза, но и от органа, в котором амилоид выпадает.
На основании анализа аминокислотного состава Леттетер (1955 г.) заключил, что амилоид - смесь двух белков: один из них близок к глобулинам сыворотки, другой - к коллагену. Белки амилоида состоят из двух фракций, обладающих различными свойствами. Фракция А (85-90%) нерастворима в воде, осаждается уксусной кислотой, после осаждения в ацетатном или фосфатном буфере при рН 3,9 - 6,4, электрофоретически близка к альфа-1- и гамма-глобулинам сыворотки, обладает антигенными свойствами. Фракция В (10-15%) растворима в воде, осаждается спиртом, обладает электрофоретической подвижностью, близкой к бета-глобулинам.
Белки плазмы в составе амилоида рассматриваются как "добавки", появление которых объясняется неспецифической адсорбцией в амилоиде многих веществ в связи с особенностями его тонкофибриллярной структуры.
Углеводы амилоида.
Полисахариды составляют 2-4% общей массы амилоида и представлены эквивалентным количеством глюкозы, галактозы, несколько меньшим количеством галактозамина и гексозамина, маннозой, фруктозой. Углеводы состоят из двух фракций: нейтральной и кислой.
Первая фракция, PAS - положительная, не дает метахромазии, обладает электрофоретической подвижностью подобно альфа-2- и гамма-глобулинам сыворотки. Ее относят к сывороточным глюкопротеидам, что подтверждается высоким содержанием в амилоиде и сыворотке крови при амилоидозе гексозамина и нейраминовой кислоты, которые входят в состав углеводных групп, связанных с сывороточными глобулинами.
Вторая фракция метахроматична, основу кислых полисахаридов составляют хондроитинсульфат (90%), гепаритинсульфат, гиалуроновая кислота, хондроитин и гепарин. Мукополисахариды имеют не гематогенное, а местное соединительнотканное происхождение, причем увеличение МПС происходит без нарушения соотношения хондроитинсульфата и гепаритинсульфата, присущего нормальной ткани. По данным Muir и Cohen (1968 г.) помимо хондроитинсульфата (50%) и гепаритинсульфата (30%) входит кератинсульфат (20%).
Фракция А связана с PAS-положительными сывороточными полисахаридами, фракция В связана с кислыми мукополисахаридами (с толуидиновым синим дает метахромазию).
Помимо белков и углеводов в амилоиде обнаружены липиды и липопротеиды, соли кальция (добавки амилоида).
Физические свойства амилоида.
Из физических свойств для амилоида характерны положительная анизотропия и дихроизм. При окраске красным Конго усиливается двойное лучепреломление. Анизотропия амилоида свидетельствует об упорядоченной молекулярной структуре его вещества. Исходя из поляризационно-оптических свойств амилоида Romhanyi полагает, что он обладает тонкофибриллярной субмикроскопической паракристаллической структурой.
Прочность белково-полисахаридных связей объясняет устойчивость амилоида к действию ферментов (например: трипсин ведет к ослаблению PAS- реакции амилоида, но не изменяет ортохроматичное окрашивание и двойное лучепреломление).
Окрашивание амилоида Конго-красным определяется конформационными особенностями белков и его фибрилл.
Ультраструктурные компоненты.
В амилоиде найдены два ультраструктурных компонента: фибриллы и периодические палочки. Фибриллы имеют диаметр 7,5 нм и длиной до 800 нм. Каждая фибрилла состоит из двух субфибрилл диаметром 2,5 нм, которые расположены параллельно на расстоянии 2,5 нм. Фибриллы имеют исчерченность, лентовидную форму. Согласно Wolman (1971 г.) параллельные фибриллы белка и нейтральных полисахаридов амилоида переплетены фибриллами из кислых мукополисахаридов. Для фибриллярного белка амилоида (F-компонент) характерно высокое содержание триптофана, дикарбоксильной кислоты, коротких цепей аминокислот и малое содержание гидроксипролина, гидроксилизина - это отличает амилоид от коллагена, ретикулина, эластина. Фибриллы представляют гетерогенную группу белков с индивидуальными особенностями, в каждом случае амилоидоза аминокислотный состав фибриллярного белка различен. Из смеси белков фибрилл амилоида выделяют белок А (AS) и белок В. Белок А (AS), по сравнению с белком В содержит больше таких аминокислот, как аргинин, аспарагин, глицин, аланин и фенилаланин; имеет специфическую антигенную структуру. Белок В, найденный в амилоиде при первичном амилоидозе, опухолевом амилоидозе имеет большую относительную молекулярную массу, по аминокислотному составу напоминает легкие цепи иммуноглобулинов. Для белков амилоида характерна складчатая упаковка полипептидных цепей (кросс-бета-конформация).
Периодические палочки составляют 5 % по отношению к фибриллам. Диаметр их 10 нм, длина до 250 нм. Они состоят из пептагональных образований диаметром 9-10 нм и шириной 2 нм, расположенных друг от друга на расстоянии 4 нм. Они отличаются от фибрилл не только по количественному составу аминокислот (преобладают глицин, лейцин, глютамин и аспарагин, отсутствует триптофан), но и по составу углеводов (низкое содержание гексозамина и уроновой кислоты, высокое содержание нейраминовой кислоты и гексоз). По сравнению с белком фибрилл периодические палочки являются более сильным антигеном. Периодические палочки (Р-компонент) соответствуют альфа-глобулину плазмы.
Патогенез амилоидоза.
Для объяснения патогенеза амилоидоза было предложено много теорий. Так, например, Gzerny (1893 год) и Шепилевский (1899) рассматривали амилоид как продукт воспаления.
Н. П. Кравков (1898 год) считал, что амилоид образуется вследствие жизнедеятельности микроорганизмов; по А. А. Максимову (1896 год) амилоидное вещество появляется в результате тканевой дезорганизации. До настоящего времени не утратили своего значения 3 теории патогенеза амилоидоза: теория диспротеиноза, иммунологическая теория и теория клеточной и локальной секреции.
Теория диспротеиноза.
Создание этой теории связывают с именем Вирхова, который рассматривал амилоид как "продукт крови". Эта теория рассматривает амилоид как продукт нарушенного белкового обмена. Основным звеном в патогенезе является диспротеинемия с накоплением в плазме грубодисперсных белковых фракций и аномальных белков (парапротеинов), которые, выйдя за пределы сосудистого русла, образуют амилоидную субстанцию.
Хорошо известно, что развитию амилоидоза предшествует состояние диспротеинемии (гиперглобулинемия, гиперфибриногенемия, гипоальбуминемия, парапротеинемия и др.). Оно наблюдается при заболеваниях, ведущих к вторичному амилоидозу (туберкулез, ревматоидный артрит, хронический остеомиелит, лимфогранулематоз, плазмоцитома), при периодической болезни и при идиопатическом амилоидозе.
Амилоид при плазмоцитоме строится из легких цепей иммуноглобулина. Установлено сходство состава и последовательности аминокислот белка фибрилл амилоида (белка В) при плазмоцитоме с N-концевыми фрагментами легких цепей иммуноглобулина.
Диспротеинемия характеризует предамилоидную стадию. Это происходит при введении микроорганизмов, токсинов, белков и протеолитических ферментов. В этой стадии в крови прогрессирует нарастание гамма-глобулинов, чему предшествует увеличение альфа- и бетта-глобулинов и соответствующих глюкопротеидов. Происходит образование аномальных белков, отсутствующих в крови контрольных животных.
Джегер (1960 г.) отметил падение уровня аномальных гамма-глобулинов сыворотки при появлении амилоидной субстанции в тканях, связав этот факт с расходом аномальных белков на построение амилоида.
Прекращение инъекций амилоидогена вело к нормализации уровня сывороточных бетта-глобулинов и увеличению гамма-глобулинов, что сопровождалось резорбцией амилоида в селезенке и почке.
Существенным звеном в патогенезе амилоидоза, которое объясняется теорией диспротеиноза, является выделение циркулирующих в плазме белков (парапротеинов) в ткани. Амилоид образуется под эндотелием и аргирофильной мембраной сосуда, что свидетельствует о проницаемости сосудистой стенки для парапротеинов.
Процессы полимеризации и деполимеризации кислых МПС основного вещества легче реализуются и легче разрушаются близ эндотелиального барьера, где они осуществляются под влиянием плазматических и тканевых гиалуронидаз. Субэндотелиальное отложение амилоида отражают изменение в ферментной системе (гиалуронидаза), приводящих к нарушению физиологических ритмов полимеризации и деполимеризации: это определяет возможность соединения плазменных парапротеинов и глюкопротеидов с фибриллярным белком и МПС основной субстанции, что ведет к образованию амилоида.
Субэндотелиальная локализация амилоида свидетельствует в пользу теории, подчеркивающей первичное значение нарушений синтеза белков организма, она объясняет избирательную органную локализацию амилоида. Выделяемые из крови продукты нарушенного синтеза белков скапливаются за эндотелиальным барьером (в органах выделения и депонирования), с чем связано частое положение при амилоидозе почек, печени, кишечника и селезенки. Однако в своих положениях эта теория слишком прямолинейна и поэтому нуждается в поправках.
Диспротеинемия неспецифична для амилоидоза и является выражением нарушенного белкового обмена, свойственного хроническим гнойно-деструктивным процессам, гипериммунизации, аутоиммунизации, наследственным метаболическим заболеваниям. Теория диспротеиноза не учитывает роль клеточных трансформаций РЭС и не вскрывает механизм развития самой диспротеинемии. Она объясняет механизм образования специфического белка амилоида при плазмоцитоме и не в состоянии ответить, где и как образуются белки и глюкопротеиды, которые являются "строительным материалом" амилоидной субстанции.
Иммунологическая теория.
Основоположником этой теории, рассматривающей образование амилоида как результат реакции антиген-антитело, считается Loeschke (1927) и Letteter (1934) . Они исходили из того, что при заболеваниях, осложняющихся амилоидозом, образуются продукты распада тканей, лейкоцитов и токсины бактерий, которые могут обладать антигенными свойствами и вести к продукции антител. Образование амилоида является результатом реакции преципитации белкового комплекса в местах продукции антител (РЭС). Letteter нашел в крови животных при амилоидозе специфические для тканевых белков преципитины, доказав тем самым их аутоиммуную природу, и установил, что амилоидоз возникает при условии плохой выработки антител, при их недостаточном содержании в крови и тканях и избыточное количество антигенов. Так частота амилоидоза при нагноительных процессах косвенно указывает на значение в амилоидогенезе тканевого распада и распада лейкоцитов и развитие аутоиммуной реакции.
Исследования Janigan и Druet (1966) показали, что амилоидогенная способность различных растворимых белковых веществ повышается параллельно усилению их антигенных свойств. Приобретение антигенных свойств сопровождается появлением способности вызывать амилоидоз, при потере антигенности вещество лишается этой способности.
Mellors и Ortega (1956) показали, что амилоид может связаться с гомологичной антигамма-глобулиновой сывороткой по типу антиген-антитело, он содержит специфические антитела к антигену - амилоидогену, при этом локализация специфических антител совпадает с фиксацией гомологичных гамма-глобулинов.
В гистосерологических исследованиях Letteter, Lachmann (1969) нашли в амилоиде комплемент и, тем самым, показали участие реакции антиген-антитело в образовании амилоида. Позже в массах амилоида были найдены фиксирующие комплемент комплексы антиген-антитело.
В 1957 г. Парнес В.А. нашла в амилоидной селезенке больных, умерших от различных заболеваний, специфические антигеннные комплексы (аутоантигены). Она считает, что аутоантигенная природа амилоидоза обусловливает развитие процессов аутоиммунизации. Павлихина Л.В. и Серов В.В. пользуясь реакцией связывания комплемента, показали постепенное накопление противоорганных антител при амилоидозе. В поздние стадии амилоидоза происходило нарастание титра противоорганных антител. Амилоид имеет определенную специфику как аутоантиген.
Зубжицкий Ю.Н. (1967) обнаружил, что при экспериментальном амилоидозе мышей комплекс чужеродный антиген-гомологичное антитело обнаруживается в органах раньше, чем выявляются отложения амилоида. При этом разрушение тканей, сопровождающееся образованием амилоида, индуцируется действием комплекса антиген-антитело в условиях избытка чужеродного антигена. Появление противоорганных антител, обнаруженных в сыворотке мышей, наступает после начала разрушения тканей в ходе образования амилоида, что позволяет признать аутоантигены этих тканей аутоиммунизированными антигенами.
Фаза образования аутоиммуного комплекса, включающего антиген казеин (7S-антитело), сменяется фазой образования амилоидной субстанции, в состав которой входит антителоподобное вещество. Это вещество проявляет свойства 19S - антител и появляется после "истощения" способности плазматических клеток продуцировать нормальные иммуноглобулины.
Главную роль в развитии амилоида играет срыв гипериммунизации: снижение функции иммунокомпетентной системы, стимуляция клеточных трансформаций, ведущих к появлению клеток (амилоидобласты), секретирующих специфический фибриллярный белок амилоида.
Рокосуев В.С. (1975 г.) считает, что любая длительная антигенная стимуляция организма сопровождается выработкой каких-то неспецифических факторов ("неспецифический фактор стимулирующий амилоидогенез", "фактор переноса"), реализующих развитие амилоидоза. Установлено, что "фактор переноса", находящийся как-будто в клетках лимфоидной ткани, стимулирует выработку амилоида у мышей с недостаточным количество инъекций иммуногена при переносе его от мышей и людей, больных амилоидозом. "Фактор переноса" имеет нуклеопротеидную природу. При этом показано, что грубая ядерная фракция, извлеченная из гомогенатов селезенки амилоидных мышей, вызвала индукцию амилоидоза. То есть существует перенос информации клетками лимфоидной системы при амилоидозе, она переносится специфическим РНК-иммунным носителем.
Предполагают, что амилоид есть результат взаимодействия амилоидостимулирующего фактора и специфического антигена. При этом исключительной становится роль клеток (амилоидобласты), продуцирующих амилоид под влиянием амилоидопродуцирующего фактора в присутствии специфических антигенов.
При амилоидозе происходит угнетение клеточного иммунитета по мере прогрессирования процесса.
Таким образом, иммунологическая теория амилоидоза уточнила и дополнила представления о роли гуморальных иммунных реакций в образовании амилоидного вещества. Суждение, что "амилоид является комплексом антиген-антитело" не может быть принято, но это не значит, что иммунные комплексы не могут входить в состав амилоида как гематогенные "добавки". Присутствие иммунных комплексов с амилоиде свидетельствует о роли реакции гуморального иммунитета в образовании амилоида.
В 1973 г. Hardt привел следующие факты: к амилоидозу приводит сильная антигенная стимуляция; клетки, "занятые" в предамилоидной и амилоидной фазе, известны как "медиаторы иммунных реакций"; в амилоидной фазе наблюдается нарушение функции иммунокомпетентных клеток, и эти нарушения коррелируют с изменениями в лимфоидных органах; амилоидные фибриллы состоят из компонентов иммуноглобулинов. Длительная антигенная стимуляция приводит к распаду Т-лимфоцитов с высвобождением веществ нуклеопротеидной природы, которые вместе со специфическим антигенами вызывают продукцию фибрилл амилоида мезенхимальными клетками, то есть нарушается гуморальный и клеточный иммунитет. Эти нарушения определяют истощение лимфоидной ткани и появление пула клеток (амилоидобластов), способных секретировать аномальный фибриллярный белок, который при соединении в тканях с другими белками и полисахаридами образует сложный гликопротеид - амилоид.
Иммунологическая теория вскрывает важное звено в патогенезе амилоидоза - изменение иммунного гомеостаза. Она позволяет объяснить экспериментальный амилоидоз и вторичный амилоидоз у человека (исключая параамилоидоз), но неприложима к идиопатическому и генетическому амилоидозу.
Мутационная теория.
Эта теория образования клона клеток-амилоидобластов может стать универсальной в объяснении патогенеза всех известных форм амилоидоза, если представить возможное разнообразие мутагенных факторов.
При вторичном амилоидозе (исключая амилоидоз при плазмоцитоме) мутации и появление клона амилоидобластов связаны с длительной антигенной стимуляцией.
При генетическом (семейном) амилоидозе речь идет о мутации гена, которая происходит в различных локусах, чем и определяется различие в составе амилоидного белка у разных людей и животных. При старческом амилоидозе имеют место подобные механизмы, так как старческий амилоидоз рассматривают как фенокопию генетического. Клеточные мутации при параамилоидозе и, возможно, при опухолевидном локальном амилоидозе обусловлены опухолевыми мутагенами. Поскольку предшественники амилоида - амилоидные фибриллы (их белки) являются слабыми иммуногенами, мутирующиеся клетки не распознаются иммунокомпетентной системой и не элиминируются. Возникает иммунологическая толерантность по отношению к белкам амилоида.
Мутационная теория амилоидогенеза позволила понять близость амилоидоза к опухолевому процессу.
Говоря о конкретных взаимоотношениях амилоидоза и опухоли выделяют три варианта сочетаний:
Амилоидоз "осложняет" опухоль, обычно рак той или иной локализации. Развивается вторичный амилоидоз, причем механизм амилоидогенеза идентична таковому при туберкулезе и хронических нагноениях.
Основные звенья:
а) деградация комплексов антиген-антитело макрофагами, активация продуктами деградации пиронинофильных и PAS - положительных клеток и синтез амилоида амилоидобластами при помощи "фактора переноса", освобождающегося из пиронинофильных клеток.
Амилоидоз при ретикулезах (плазмоцитоме, ретикулосаркоме, лимфогранулематозе, болезнь Вальденштрема).
Развивается генерализованный амилоидоз, однако развитие его в этих случаях связывается с опухолевой пролиферацией клеток типа ретикулярных, плазмоцитов и лимфоцитов. При этом имеются факты, что амилоид при гемобластозах (парапротеинемических) прямо или опосредованно являются продуктом опухолевых клеток. в норме элементы линии "ретикулярная клетка - плазмоцит" синтезируют иммуноглобулины, а в процессе опухолевой трансформации - миеломные белки, чаще L - цепи иммуноглобулинов, из которых строится амилоид. предполагают, что легкие цепи иммуноглобулинов захватываются макрофагами, которые перерабатывают L - цепи и строят из этих фрагментов амилоид. L - цепи являются филогенетически более примитивными вариантами "предшественников иммуноглобулинов".
Амилоидоз стромы доброкачественных или злокачественных опухолей. Чаще это опухоли эндокринных органов или представители паракринной системы: медуллярный рак щитовидной железы, инсулома, карциноиды, кожные и одонтогенные опухоли - опухоль Тередберга. В этом случае в образовании амилоида участвуют клетки эпителиальные опухолевые клетки. В медуллярном раке щитовидной железы амилоид продуцируется парафолликулярными С-клетками, которые в норме производят кальцитонин. В инсуломах В-клетки производят амилоид или инсулин, для белковой молекулы которого, так же как для амилоида и кератина, характерно обилие дисульфидных связей. По-видимому, с этой "ошибочной" продукцией В-клетками амилоида вместо инсулина связан старческий амилоидоз островков Лангерганса (компонент триады Шварца), а с продукцией клетками эпидермиса вместо кератина амилоида - локальный кожный амилоидоз.
Связь между раковой клеткой и амилоидом при эндокринных полипептидных опухолях ("апудомах"), которые в норме продуцируют полипептиды, а в результате опухолевой трансформации наряду с другими проявляющимися признаками опухолевого прогрессии начинают синтезировать аномальный белок- "апуд- амилоид".
Таким образом, амилоид является продуктом клетки, претерпевшей опухолевую эволюцию.
Между амилоидогенезом и онкогенезом можно найти много общего. Так, признак прогрессии в амилоидозе представлен безудержным размножением недифференцированных клеток РЭС. Бесконтрольно прогрессирующую продукцию аномального белка амилоида пролиферирующими клетками особого типа можно рассмотреть как вторичный признак прогрессии (анаплазия), которая развивается автономно. Предполагают, что клетками, способными к опухолевой эволюции, является "пул" дремлющих примитивно устроенных клеток, подверженных соматическим мутациям (клетки, синтезирующие амилоидные фибриллы): это амилоидобласты, являющиеся производными камбиальных клеточных элементов мезенхимального происхождения.
Взаимоотношения амилоидоза и опухоли с позиций нарушения ферментативного баланса клетки (ферментопатия). В основе действия онкогенного фактора (вирус, соматическая мутация), по-видимому, лежит ферментопатия, главным образом РНК). Если понимать процесс "дисдифференцировки" и опухолевую эволюцию клеток как ферментопатию, обусловленную нарушением работы ферментов, связанных с обменом нуклеиновых кислот, то "дисдифференцировка" клеток с образованием амилоидобластов и синтезом амилоидного фибриллярного белка следует связать с нарушением работы ферментов, контролирующих синтез фибриллярных белков. Считают, что первичный дефект при амилоидозе заключается в нарушении синтеза нуклеопротеидов, что свидетельствует о близости тонких механизмов онко- и амилоидогенеза. В основе извращений функции клетки, ведущих к синтезу аномального фибриллярного белка, лежит мутация гена, отвечающего за выработку белка.
Генетический амилоидоз, развивающийся в связи с мутацией гена, этот амилоидоз относят к полигенным заболеваниям, развитие которых зависит от нарушения не одного фермента, а их комплекса.
Амилоид образуется в тех опухолях, которые относятся к генетически зависимым: медуллярный рак щитовидной железы, множественный эндогенный аденоматоз, опухоли при иммунодефицитных заболеваниях.
Гуморальные нарушения (диспротеинемия, извращения иммуногенеза), как и клеточные (синтезирующие белок амилоида) могут возникнуть после того или иного заболевания или быть его выражением (опухоль), могут явиться проявлением возрастаюших изменений обмена тканевой структуры или быть наследственными, отражая индивидуальную структуру белкового обмена.
Таким образом, мутационный механизм ведет к появлению амилоидобластов, секретирующих специфичный белок амилоида.
Морфогенез амилоидоза.
Морфогенез амилоидоза складывается из следующих звеньев:
Клеточные трансформации РЭС с появлением клона клеток, способных к синтезу фибриллярного белка амилоида.
Синтез амилоидобластами основного компонента амилоида - фибриллярного белка (фибриллы).
Агрегация фибрилл с образованием "каркаса" амилоидной субстанции.
Соединение агрегированных фибрилл с белками и глюкопротеидами плазмы, а также с кислыми МПС и образование сложного глюкопротеида - амилоида.
Клеточные трансформации РЭС составляют сущность предамилоидной стадии, которая изучена при вторичном амилоидозе.
Предамилоидная стадия - пиронинофильная стадия, характеризующаяся плазматизацией органов РЭС (селезенки, костного мозга, лимфатических узлов и печени) и в других органах (клубочки почек, перибронхиальная соединительная ткань, строма миокарда). Богатые РНК пиронинофильные клетки отличаются высокой активностью окислительно-восстановительных ферментов (глюкозо-6-фосфатдегидрогеназа). По мере снижения пиронинофилии в клетках увеличивается количество гидролитических ферментов (PAS - положительные клетки).
Клеточные трансформации РЭС в предамилоидной стадии обязаны двум классам клеток - плазмоцитам и лимфоцитоподобным.
В селезенке наряду с плазматическими клетками, которые в этой стадии преобладают, увеличивается количество ретикулярных клеток с гиперплазированной гранулярной ЭПС и множеством полисом.
В печени плазматические клетки и лимфоцитоподобные клетки постоянно обнаруживаются в синусоидах.
В почках плазматические и лимфоцитоподобные клетки видны в межканальцевой строме.
Таким образом, клеточные трансформации в предамилоидной стадии обязаны гиперплазии плазматическим и лимфоцитоподобным клеткам.
Синтез фибриллярного белка амилоида клетками мезенхимального происхождения.
Большинство авторов считает причастным к амилоидогенезу 2 класса фибробластов:
фибробласты, близкие к ретикулярной клетке, ответственные за продукцию белка ретикулина;
фибробласты, продуцирующие белок соединительной ткани - тропоколлаген.
Помимо фибробластов, синтез фибрилл амилоида связывают с эндотелиальными клетками.
Источниками амилоидобластов являются:
В селезенке - ретикулярные и плазматические клетки.
В печени - купферовы клетки, способные при определенных условиях синтезировать и другой фибриллярный белок - тропоколлаген.
В почках (их клубочках) отложение фибрилл амилоида чаще обнаруживается в мезангиуме; амилоидобластами становятся мезангиальные клетки, которые в норме выполняют роль модифицированных фибробластов, секретирующих тропоколлаген базальной мембраны капилляров клубочка, а также и роль макрофага.
Агрегация фибрилл амилоида с образованием "каркаса" амилоидной субстанции зависит от гуморальных и тканевых (клеточных факторов).
Среди гуморальных факторов имеют значение дисульфидные связи и сульфгидрильные группы амилоида, участвующих в агрегации белков в стабильные макроструктуры.
При образовании фибрилл амилоида, который является аномальным белком, закономерны клеточные реакции резорбции - фибриллоклазии.
Возникает как бы единоборство синтеза фибриллярного белка амилоида и его резорбции, единоборство амилоидобластов и амилоидокластов. Оно заканчивается в пользу синтеза фибриллярного белка амилоида, что объясняется развитием толерантности к амилоидному белку. Толерантность к белку фибрилл амилоида, выражением которого является недостаточная и несостоятельная амилоидоклазия, может рассматриваться как одна из главных причин накопления амилоидного вещества в тканях.
Амилоидокласты ультраструктурно отличаются от амилоидобластов. В их цитоплазме отсутствует гранулярная ЭПС, рибосомы, митохондрии, редко появляется гладкая ЭПС. Цитоплазма заполнена включениями с одноконтурной мембраной, содержащими амилоидные фибриллы. Включения отражают этапы "переваривания" фибриллярного белка амилоида (фибриллы характеризуются разным расположением, фаголизосомы, липидные капли).
В селезенке роль амилоидокластов выполняют свободные и фиксированные макрофаги гематогенного происхождения.
В печени - купферовы клетки.
Таким образом, агрегация фибрилл амилоида связана с двумя сопряженными разнонаправленными процессами - синтезом фибрилл и их резорбцией.
Преобладание синтетических процессов над резорбтивными (амилоидобластов над амилоидокластами) в силу восстановления иммунологической толерантности к белку амилоида определяет возможность образования фибриллярного каркаса амилоидной субстанции.
Соединение фибрилл амилоида с белками и гликопротеидами плазмы и кислыми МПС ткани представляет заключительный этап формирования амилоидного вещества.
Образование амилоида происходит вне клеток, в тесной связи с волокнами соединительной ткани - ретикулярными, либо коллагеновыми. Отношение амилоидной субстанции к разным волокнам связано с участием различных клеток (плазматические, ретикулярные клетки или фибробласты) в построении фибриллярного белка амилоида.
Существует два вида амилоида в зависимости от отношения его к фибриллярным структурам соединительной ткани - периретикулярный и периколлагеновый.
Для периретикулярного амилоида, который выпадает по ходу содержащих ретикулин мембран сосудов и желез, а также по ходу ретикулярой стромы паренхиматозных органов, типично поражение селезенки, печени, почек, надпочечников, кишечника, интимы сосудов мелкого и среднего калибра.
Для периколлагенового амилоида, который образуется по ходу коллагеновых волокон, характерно преимущественное поражение адвентиции сосудов крупного и среднего калибра, стромы миокарда, поперечно-полосатой и гладкой мускулатуры, нервов, кожи (мезенхимальный амилоидоз).
На заключительном этапе амилоидогенеза увеличивается сосудистая проницаемость, она облегчает соединение фибриллярного белка амилоида и МПС ткани с белками и глюкопротеидами плазмы крови и определяют присутствие в амилоиде гематогенных "добавок".
Прогрессирующий амилоидоз ведет к функциональной недостаточности органа, ведет к атрофии его паренхиматозных элементов и склерозу. Орган увеличивается в объеме, становится плотным и ломким, имеет восковидный или сальный вид на разрезе; в финале развивается амилоидное сморщивание.
Амилоидоз может быть генерализованным (общий, распространенный амилоидоз) или местным (локальный).
Классификация амилоидоза.
В 1935 г. Reimann с соавторами предложили классификацию амилоидоза, основанную на клинико-морфологических особенностях его видов.
В 1929 г. Lubarsh писал, что первичный амилоидоз отличается от вторичного отсутствием предшествующего или сопутствующего "причинного" заболевания, более частым поражением мезодермальных тканей, по сравнению с паренхимными (сердечно-сосудистая система, пищеварительный тракт, поперечно-полосатые и гладкие мышцы кожи). Причины первичного амилоидоза неизвестны, хотя учитывается подобие типов наследственного и первичного амилоидоза.
Таблица 1.
Классификация амилоидоза Heller и соав. (1984).
| Форма | Вид отложений | амилоидоза |
| амилоидоза | периретикулярный | периколлагеновый |
Генетический (наследственный) | Семейная средиземноморская лихорадка (периодическая болезнь). Амилоидоз с лихорадкой, крапивницей и глухотой (форма Muckle и Wells) | Нейропатический амилоидоз с преимущественным поражением нижних (Andrade, Horta) или верхних (Rukavina) конечностей. Кардиопатический амилоидоз (Frederiksen). |
| Приобретенный (вторичный) | Ассоциированный с хроническими инфекциями, коллагеновыми болезнями, злокачественными опухолями. | Ассоциированный с множественной миеломой |
| Идиопатический (первичный) | Нефропатический | Классический (системный). Нейропатический. Кардиопатический. Локальный. |
Причинами вторичного амилоидоза может быть туберкулез, который конкурирует с гнойно-деструктивными процессами в легких, а также велика роль в возникновении амилоидоза ревматоидного артрита и остеомиелита. Амилоидоз при миеломной болезни (плазмоцитома) выделен в самостоятельную форму, поскольку он занимает промежуточное положение между вторичным и первичным: так как при вторичном имеется "причинное" заболевание - миелома, однако характер распределения (сосуд, локализация) и химико-тинкториальные свойства амилоидного вещества ближе к первичной форме амилоидоза.
Некоторые исследователи полагают, что механизм образования амилоида при миеломной болезни в отличие от типичного амилоидоза более простой и сводится к диффузии в ткани микромолекулярного сывороточного белка Бенс-Джонса, который вступает в реакцию "белок- белок" или "белок- полисахариды", что ведет к преципитации нерастворимового параамилоида.
Однако в настояшее время имеются доказательства в пользу того, что при плазмоцитоме фибриллярный белок амилоида строится из лёгких цепей иммуноглобулина, секретируемого миеломной клеткой.Таков же механизм образования при парапротеинемических гемобластозах.
В 1961 году Бриггс восполнил классификацию амилоидоза:
1) Первичный амилоидоз (отсутствие предраспологающего заболевания)
а) генерализованный
б) семейный
в) респираторного тракта (опухолевидный, узловатый и диффузный)
2) Вторичный амилоидоз (наличие предраспологающего заболевания)
3) Старческий амилоидоз (сердца)
4) Амилоидоз при миеломной болезни
5)Опухолевидный локализованный амилоидоз (кроме амилоидоза респираторного тракта).
Гафни в 1964 году свёл клинико-морфологические особенности наследственного амилоидоза к трём типам:
1) нефропатический, проявляющейся нефротическим синдромом с уремией в финале заболевания
2) нейропатический, характеризующийся полиневритом с мышечной атрофией, импотенцией, кишечными расстройствами и кахексией
3) кардиопатический, характеризующийся сердечной недостаточностью.
Исходя из материалов последних лет по изучению амилоидоза,
Серов В.В. предлагает классификационную схему амилоидоза, которая учитывает:
1) Форму амилоидоза в зависимости от причинного фактора;
2) Возможный патогенетический механизм;
3) Клинический тип амилоидоза в зависимости от преобладания поражений того или иного органа или системы;
4) Морфогенетический вид в связи с особенностями периретикулярного или периколлагенового расположения амилоидной субстанции;
5) Клинико-морфологический вариант амилоидоза-паренхиматозный или мезенхимальный.
Существуют ряд общих признаков, которые объединяют амилоидоз в целом. Это, во-первых, диспротеинемия,которая является выражением нарушенного обмена, а возможно, и особенностей процесса обновления белков тела; во-вторых, единая роль трансформации клеток РЭС в возникновении фибриллярной структуры амилоида, независимо от того, является ли эта трансформация реактивной, неопластической или она обусловлена генетически; в-третьих, предшествующиепоявлению амилоида субмикроскопические изменения элементов системы соединительной ткани; в-четвёртых, единая субмикроскопическая структура амилоида.
Таблица № 2. Классификация амилоидоза.
| Форма 1 | Основной механизм развития 2 | Клинический тип 3 | Морфологический вид 4 | Клинико-морфологический вариант 4 |
| Идиопатический (первичный) | Неизвестен | Системный Кардиопатический Нейропатический Нефропатический Энтеропатический Гепатопатический | Периколлагеновый >> Периретикулярный >> >> | Мезенхимальный >> >> Паренхиматозный >> >> |
| Наследственный (генетический) | ||||
| 1) периодическая болезнь (семейная средиземноморская лихорадка ) | Нефропатический | >> | >> | |
| 2) семейный амилоидоз с лихорадкой, крапивницей, глухотой | Генетический дефект синтеза фибриллярных белков тела (наследственная ферментопатия) | >> | >> | >> |
| 3) семейный амилоидоз с аллергическими проявлениями, лихорадкой и нефропатией) | >> | >> | >> | |
| 4) семейный нейропатический амилоидоз | Нейропатический | Периколлагеновый | Мезенхимальный | |
| тип I, португальский | ||||
| тип I I | ||||
| 1 | 2 | 3 | 4 | 5 |
| тип III | ||||
| тип IV | ||||
| 5) семейный кардиопатический амилоидоз | Кардиопатический | >> | >> | |
| Приобретенный (вторичный) 1) амилоидоз как осложнений хронических инфекций, коллагеновых болезней и злокачественных опухолей | Нарушение иммунологического гомеостаза | Нефропатический Эпинефропатический Гепатопатический Смешанный | Периретикулярный >> Периколлагеновый | Паренхиматозный >> Мезенхимальный |
| 2) параамилоидоз | Неопластическая трансформация клеток белково-синтетической системы | |||
| Старческий амилоидоз | Инволютивные нарушения обмена белка. возрастная брадитрофия тканей | Кардиопатический | >> | >> |
| Локальный опухолевидный | Неизвестен |
Похожие работы
... периодической болезнью с амилоидозом, вскоре после назначения КС появились признаки почечной недостаточности. Обычно выделяют, хотя весьма условно, несколько стадий в развитии амилоидоза почек: латентную (бессимптомную), альбуминурическую, отечную и терминальную, протеинурическую, отечно-гипотоническую и азотемическую. О существовании скрытой (бессимптомной) стадии сообщает Б. М. Ковалив (1970 ...
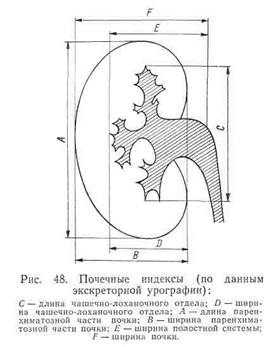

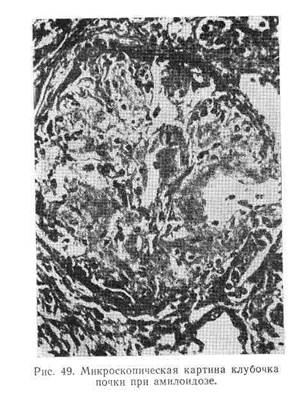
... биопсии, так как в связи с гнездностью процесса полученные отрицательные данные не исключают наличия пиелонефрита. Дифференциальный диагноз. Хронический пиелонефрит следует дифференцировать с хроническим гломерулонефритом, амилоидозом почек, диабетическим гломерулосклерозом, гипертонической болезнью. Нельзя забывать о возможности комбинированных поражений (гломеруло- и пиелонефрит, диабетический ...
... выпадают по ходу коллагенновых волокон. Чаще всего поражаются органы мезенхимального происхождения. При идиопатической форме поражаются сердце, нервы, кишечник. При наследственном или семейном амилоидозе поражаются симпатические нервные ганглии, а также паренхиматозные органы - почки. Очень характерна так называемая периодическая болезнь, которая встречается у лиц наиболее древних национальностей: ...
... Легочная гипертензия I ст. Атеросклероз аорты, мозговых артерий. Артериальная гипертензия 3 ст. риск 4. Гипертоническая ангиопатия сетчатки. Фоновое: Экзогенно-конституциональное ожирение 2 ст. Сопутствующие: - Осложнения: НК IIА ФК3 Диагноз AL- амилоидоз поставлен на основании жалоб больного на отёки на нижних и верхних конечностях, лице, слабость, снижение работоспособности, по данным ОАК ...
0 комментариев