Навигация
Обработка культуры клеток алкалоидом колхицином, который ведет к накоплению делящихся клеток на стадии метафазы;
73959
знаков
1
таблица
3
изображения
1. Обработка культуры клеток алкалоидом колхицином, который ведет к накоплению делящихся клеток на стадии метафазы;
2. Обработка клеток слабыми растворами солей, вызывающими набухание, расправление хромосом, что облегчает их исследование.
В 1956 г. шведские цитологи Дж. Тийо и А. Леван изготовили культуры клеток из тканей легких, взятых у абортированных человеческих эмбрионов и, используя усовершенствованную методику обработки клеток, получили необычайно четкие препараты, в которых ясно было видно 46 хромосом.[5]
Несколькими месяцами позднее Ч. Форд и Дж. Хаммертон в Англии установили, что диплоидные предшественники половых клеток в семенниках мужчин (сперматогонии) также имеют по 46 хромосом, а гаплоидные (сперматоциты 1-го деления) — по 23 хромосомы.
После этого были изучены многие клетки из разных органов и тканей человека и везде нормальное число хромосом оказалось равным 46.
Женский кариотип отличается от мужского только одной половой хромосомой. Остальные 22 пары одинаковы у мужчин и женщин. Эти 22 пары хромосом называются аутосомами. Нормальный кариотип состоит из 44 аутосом (22 пары) и двух половых хромосом — XX у женщин и XY у мужчин, т. е. женский кариотип имеет две большие половые хромосомы, а мужской — одну большую и одну маленькую.
В половых клетках человека находится одинарный (гаплоидный) набор хромосом — 23, а в соматических клетках — двойной (диплоидный) набор — 46. Эти открытия стимулировали дальнейшее изучение хромосом. Были разработаны методы исследования хромосом в культуре лимфоцитов периферической крови и на других объектах. В настоящее время хромосомы относительно легко исследуют в лимфоцитах периферической крови. Венозную кровь помещают в специальную питательную среду, добавляют фитогемаглютинин, который стимулирует клетки к делению, и помещают на 72 ч. в термостат. За 6 ч. до конца инкубации сюда добавляют колхицин, который задерживает процесс деления клеток на стадии метафазной пластинки. Затем культуру помещают в гипотонический раствор NaCl, в котором клетки набухают, что приводит к легкому разрыву оболочек ядра и переходу хромосом в цитоплазму. После этого препараты окрашивают ядерными красителями, в частности ацетоорсеином, и рассматривают их в световом микроскопе с иммерсией.
Под микроскопом учитывают общее количество хромосом, фотографируют их, затем из фото вырезают ножницами каждую хромосому и наклеивают на чистый лист бумаги в ряд, начиная от самой большой (первой) хромосомы и кончая самой маленькой (двадцать второй) и половой Y-хромосомой. Люминесцентная методика позволяет быстро и просто проводить массовые исследования с целью выявления больных с различными типами хромосомных аномалий. Совокупность количественных (число хромосом и их размеры) и качественных (морфология хромосом) признаков диплоидного набора единичной клетки обозначается термином «кариотип». Строение хромосом изменяется в зависимости от стадии деления клеток (профазы, метафазы, анафазы, телофазы).
Уже в профазе митоза видно, что хромосома образована двумя взаимно переплетающимися нитями одинакового диаметра — хроматидами. В метафазе хромосома уже спирализована, и две ее хроматиды ложатся параллельно, разделенные узкой щелью. Каждая хроматида состоит из двух полухроматид. В результате митоза хроматиды материнской хромосомы становятся сестринскими хромосомами, а полухроматиды — их хроматидами. В основе хроматид лежат хромонемы — так называют более тонкие нити ДНП, состоящие из белка и нуклеиновых кислот.
В интерфазе (промежуток между двумя делениями клеток) хроматин тесно связан с ядерными мембранами и ядерным белковым матриксом. Он образует также большие участки деспирализованных нитей ДНП. Затем постепенно хроматин спирализуется, образуя типичные метафазные 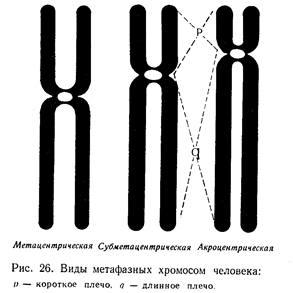 хромосомы. Размеры их варьируют от 2 до 10 микрон.
хромосомы. Размеры их варьируют от 2 до 10 микрон.
В настоящее время интенсивно исследуются структурные особенности аутосом и половых хромосом (на клетках костного мозга, лимфоцитах, фибробластах, клетках кожи, регенерирующей печени).
![]() В хромосомах выявлены структуры, названные хромомерами. Хромомер — это спирализованный участок хромонемы. Промежутки между хромомерами представлены хромонемными нитями. Расположение хромомеров на каждой хромосоме строго фиксировано, наследственно детерминировано.
В хромосомах выявлены структуры, названные хромомерами. Хромомер — это спирализованный участок хромонемы. Промежутки между хромомерами представлены хромонемными нитями. Расположение хромомеров на каждой хромосоме строго фиксировано, наследственно детерминировано.
Хромомер — сравнительно крупная генетическая единица, сравнимая по длине с хромосомой кишечной палочки. Строение и функция хромомера — основная загадка современной генетики. Предполагают, что некоторые хромомеры — это один генетический локус, где есть один структурный ген и много генов регуляторных. Возможно, в других хромомерах располагается несколько структурных генов.
Хромонемы и хромомеры окружены неокрашивающимся веществом — матриксом. Полагают, что матрикс содержит дезоксирибонуклеиновую и рибонуклеиновую кислоты, белки.
Определенные участки хромосом образуют ядрышки. Ядрышки — это более или менее деспирализованные участки хромосом, окруженные продуктами деятельности генов (рибосомы, частицы РНК и т. п.). Здесь идет синтез рибосомальной РНК, а также осуществляются определенные этапы формирования рибосом. В нем синтезируется большая часть РНК клетки.
В метафазной хромосоме различают еще несколько образований: центромеру, два плеча хромосомы, теломеры и спутник.
Центромерный (meros — по-гречески, часть) участок хромосомы — это неокрашивающийся разрыв в хромосоме, видимый на препарате хромосом. Центромера содержит 2—3 пары хромомер, имеет сложное строение. Предполагают, что она направляет движение хромосомы в митозе. К центромерам прикрепляются нити веретена.
Теломеры — специальные структуры на концах хромосом — также имеют сложное строение. В их состав входит несколько хромомер. Теломеры предотвращают концевое присоединение метафазных хромосом друг к другу. Отсутствие теломеров делает хромосому «липкой» — она легко присоединяется к другим фрагментам хромосом.
Одни участки хромосомы называются эухроматиновыми, другие — гетерохроматиновыми. Эухроматиновые районы хромосом — это генетически активные участки, они содержат основной комплекс функционирующих генов ядер. Потеря даже мельчайшего фрагмента эухроматина может вызвать гибель организма. Гетерохроматиновые районы хромосом — обычно сильно спирализованы и, как правило, генетически мало активны. В гетерохроматине находится ядрышковый организатор. Потеря даже значительной части гетерохроматина часто не приводит организм к гибели. Гетерохроматиновые участки хромосомы реплицируются позднее, чем эухроматиновые. Следует помнить, что эухроматин и гетерохроматин — это не вещество, а функциональное состояние хромосомы.
Если расположить фотографии гомологичных хромосом по мере возрастания их размеров, то можно получить так называемую идиограмму кариотипа. Таким образом, идиограмма — это графическое изображение хромосом. На идиограмме пары гомологов располагаются рядами в порядке убывающего размера.
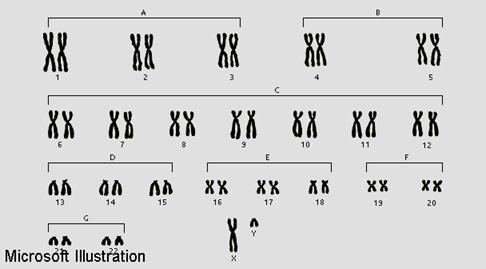
У человека на идиограмме среди 46 хромосом различают три типа хромосом в зависимости от положения в хромосоме центромер:
1. Метацентрические — центромера занимает центральное положение в хромосоме, оба плеча хромосомы имеют почти одинаковую длину;
2. Субметацентрические — центромера располагается ближе к одному концу хромосомы, в результате чего плечи хромосомы разной длины.
| Классификация хромосом человека по размеру и расположению центромера | ||
| Группа хромосом | Номер по кариотипу | Характеристика хромосом |
| А(1) | 1,2,3 | 1 и 3 почти метацентрические и 2—крупная субметацентрическая |
| В (11) | 4,5 | крупные субакроцентрические |
| С (III) | 6—12 | средние субметацентрические |
| A(lV) | 13—15 | средние акроцентрические |
| E(V) | 16-18 | мелкие субметацентрические |
| F(VI) | 19—20 | самые мелкие мегацентрические |
| G(VII) | 21—22 | самые мелкие акроцентрические |
| Х-хромосома (относится к III группе | 23 | средняя почти метацентрическая |
| Y-хромосома | 23 | мелкая акроцентрическая |
3. Акроцентрические — центромера находится у конца хромосомы. Одно плечо очень короткое, другое длинное. Хромосомы не очень легко отличать одну от другой. Цитогенетики с целью унификации методов идентификации хромосом на конференции в 1960 г. в г. Денвере (США) предложили классификацию, учитывающую величину хромосом и расположения центромер. Патау в том же году дополнил эту классификацию и предложил разделить хромосомы на 7 групп. Согласно этой классификации, к первой группе А относятся крупные 1, 2 и 3 суб- и акроцентрические хромосомы. Ко второй группе В — крупные Субметацентрические пары 4—5. К третьей группе С относятся средние субакроцентрические (6—12 пары) и Х-хромосома, которая по величине находится между 6 и 7 хромосомами. К группе Д (четвертой) относятся средние акроцентрические хромосомы (13, 14 и 15 пары). К группе Е (пятой)— мелкие Субметацентрические хромосомы (16, 17 и 18 пары). К группе F (шестой) мелкие метацентрические (19 и 20 пары), а к группе G (седьмой) — самые мелкие акроцентрические хромосомы (21 и 22 пары) и мелкая акроцентрическая половая Y-хромосома (табл. 4).
Существуют и другие классификации хромосом (Лондонская, Парижская, Чикагская), в которых развиты, конкретизированы и дополнены положения Денверской классификации, что в конечном итоге облегчает идентификацию и обозначение каждой из хромосом человека и их частей.
Акроцентрические хромосомы IV группы (Д, 13—15 пары) и группы VII (G, 21—22 пары) на коротком плече несут маленькие дополнительные структуры, так называемые сателлиты. В некоторых случаях эти сателлиты являются причиной сцепления хромосом между собой при делении клеток в мейозе, вследствие чего происходит неравномерное распределение хромосом. В одной половой клетке оказывается 22 хромосомы, а в другой — 24. Так возникают моносомии и трисомии по той или иной паре хромосом. Фрагмент одной хромосомы может присоединиться к хромосоме другой группы (например, фрагмент 21 или 22 присоединяется к 13 или 15). Так возникает транслокация. Трисомия 21-й хромосомы или транслокация ее фрагмента являются причиной болезни Дауна.
Внутри семи этих групп хромосом на основании лишь внешних различий, видимых в простой микроскоп, провести идентификацию хромосом почти невозможно. Но при обработке хромосом акрихини притом и при помощи ряда других методов окраски их можно идентифицировать. Известны различные
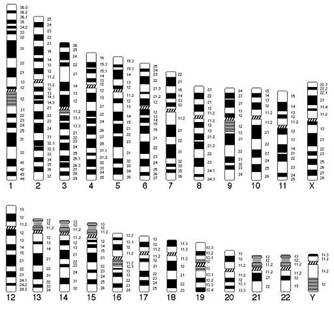
способы дифференциальной окраски хромосом по Q-, G-, С-технике (А. Ф.Захаров, 1973) (рис. 27). Назовем некоторые методы идентификации индивидуальных хромосом человека. Широко применяются различные модификации так называемого метода Q. Например, метод QF — с использованием флюорохромов; метод QFQ — с использованием акрихина; метод QFH — с использованием специального красителя фирмы «Хекст» № 33258, выявляющего повторяющиеся последовательности нуклеотидов в ДНК хромосом (сателлитную ДНК и т. п.). Мощным средством изучения и индивидуальной характеристики хромосом являются модификации трипсинового метода GT. Назовем, например, GTG-метод, включающий обработку хромосом трипсином и окраску красителем Гимза, GTL-метод (обработка трипсином и окраска по Лейтману).
Известны методы с обработкой хромосом ацетатными солями и красителем Гимза, методы с использованием гидроокиси бария, акридиноранжа и другие.
ДНК хромосом выявляется при помощи реакции Фельгена, окраски метиловым зеленым, акридиноранжем, красителем № 33258 фирмы «Хекст». Акридиноранжевый краситель с ДНК однонитчатой образует димерные ассоциаты и дает красную люминесценцию, с двунитчатой спиральной ДНК образует одномерные ассоциаты и люминесцирует зеленым светом.
Измеряя интенсивность красной люминесценции, можно судить о количестве свободных мест в ДНП и хроматине, а отношение зеленая — красная люминесценция — о функциональной активности хромосом.
Гистоны и кислые белки хромосом выявляются при различных рН окраской бромфенодовым синим, зеленым прочным, серебрением, иммунолюминесцентным методом, РНК — окраской галлюцианиновыми квасцами, красителем фирмы «Хекст» № 1, акридиноранжем при нагревании до 60°.
Широко применяются электронная микроскопия, гистоавторадиография и ряд других методов.
В 1969 г. шведский биолог Т. Касперссон и его сотрудники показали, что хромосомы, окрашенные горчичным акрихином и освещенные под микроскопом Наиболее длинноволновой частью ультрафиолетового спектра, начинают люминесцировать, причем одни участки хромосом светятся ярче, другие слабее. Причина этого — разный химический состав поверхности хромосомы. В последующие годы исследователи обнаружили, что концы Y-хромосомы человека светятся ярче любой другой хромосомы человека, поэтому Y-хромосому легко заметить на препарате.
Акрихиниприт преимущественно связывается с ГЦ-парами ДНК. Флюоресцируют отдельные диски гетерохроматиновых участков. Удаляют ДНК — свечение исчезает. Составлены карты флюоресцирующих хромосом. Из 27 видов млекопитающих только у человека, шимпанзе, гориллы и орангутанга светятся Y-хромосомы. Свечение связано с повторами генов, которые появились в эволюции 20 млн. лет назад.
Итак, в норме в соматических клетках человека находится 46 хромосом (23 пары), а в половых — 23 хромосомы, по одной хромосоме каждой пары. При слиянии сперматозоида и яйцеклетки в зиготе количество хромосом удваивается. Таким образом, каждая соматическая клетка организма человека содержит один набор отцовских хромосом и один набор материнских хромосом. Если у человека 46 хромосом, то у различных обезьян число хромосом равно 34, 42, 44, 54, 60, 66.
При действии ультразвука или высокого давления можно добиться разрыва нитей ДНК, которые входят в состав хромосомы, на отдельные фрагменты. Подогревая растворы ДНК до температуры 80—100°,
можно вызвать денатурацию ДНК, расхождение двух составляющих ее нитей. При определенных условиях разъединенные нити ДНК могут снова реассоциировать в устойчивую двунитчатую молекулу ДНК (реассоциация или ренатурация ДНК). Денатурацию и ренатурацию ДНК можно получить и на препаратах фиксированных хромосом, обрабатывая их соответствующим образом. Если после этого хромосомы окрасить красителем Гимза, то в них выявляется четкая поперечная исчерченность, состоящая из светлых и темных полос. Расположение этих полос в каждой хромосоме разное. Таким образом, по «Гимза-дискам» можно также идентифицировать каждую из 23 пар хромосом.
Этими и другими методиками, особенно гибридизацией соматических клеток различных животных и человека, пользуются для картирования хромосом, т. е. для определения положения разных генов в той или иной хромосоме. В настоящее время в аутосомах и половых хромосомах человека картировано около 200 генов.
На конец 1975 г. было локализовано следующее количество генов в различных хромосомах человека (А. Ф. Захаров, 1977): 1 хромосома — 24 гена; 2 хромосомы — 10, 3—2, 4—3, 5—3, 6—14, 7—4, 8—1, 9—8, 10—5, 11—4, 12—10, 13—3, 14—3, 15—6, 16—4, 17—14, 18—1, 19—4, 20—3, 21—4, 22—1; Y-хромосома — 2; Х-хромосома — 95 генов.
Глава 4. Половой хроматин.В 1949 г. М. Барр и Ч. Бертрам, изучая нейроны кошки, обратили внимание на то, что в интерфазном ядре клетки содержится интенсивно окрашиваемое тельце, причем оно присутствует только в ядрах клеток самок и отсутствует у самцов. Оно было найдено у многих животных и всегда только у особей женского пола. Это тельце получило название полового хроматина, или тельца Барра. У ряда позвоночных и у человека оно появляется в раннем онтогенезе на стадии гаструлы, но раньше развития гонад (половых желез). На локализацию, форму и структуру полового хроматина не влияют половые гормоны, следовательно, он не является вторичным половым признаком. Между числом телец полового хроматина и числом X-хромосом в ядре имеется прямая связь. Половой хроматин в интерфазных ядрах обусловлен спирализацией одной из Х-хромосом, инактивация которой является механизмом, выравнивающим баланс генов половых хромосом в клетках самцов и самок (т. е. это один из механизмов дозовой компенсации генов).[6]
В 1961 г. несколько исследователей одновременно высказали предположения, что одна из Х-хромосом у нормальных женщин относительно не активна в генетическом отношении. В 1961 году английская исследовательница М. Лайон выдвинула гипотезу о механизмах инактивации одной из Х-хромосом клеток женского организма. Основные положения этой гипотезы следующие:
1. Одна из двух Х-хромосом клеток женщины неактивна.
2. Неактивная хромосома может быть отцовского или материнского организма.
3. Инактивация происходит в раннем эмбриогенезе и сохраняется во время дальнейшего размножения и развития клеточной линии. Этот процесс инактивации Х-хромосомы в ряду поколений обратим:
XX* ->- УХ -> XX* и т. д. (здесь звездочкой обозначена спирали-зованная Х-хромосома). Такой тип обратимых изменений генетического материала португальский генетик Серра предложил называть трепцией (от греч. treptos — изменение).
Спирализованная Х-хромосома в клетке образует половой хроматин или тельце Барра. Если у женщин в ядре клетки несколько Х-хромосом, то в клетках несколько телец Барра, активной остается лишь одна Х-хромосома. Х-хромосома инактивируется не вся, часть короткого плеча остается генетически активной. Инактивация Х-хромосомы в определенной мере зависит от стадии клеточного цикла и физиологического состояния организма. По наличию лишнего или отсутствию тельца Барра можно диагносцировать некоторые виды наследственных заболеваний (например, синдром Клайнфельтера, синдром Шерешевского — Тернера). Клетки, не содержащие половой хроматин (хроматин-отрицательные клетки), обнаруживаются у индивидуумов, имеющих набор хромосом 45, ХО (синдром Шерешевского — Тернера);
46, XY (нормальные мужчины); 47, XYY (синдром Клайнфельтера с двумя Y-хромосомами). Обычно в клетках нормального мужского организма встречается некоторое количество псевдотелец Барра (конденсированных участков аутосом) и спирализованных Y-хромосом, поэтому при диагностике различных хромосомных заболеваний необходимо уметь отличать эти образования от типичного полового хроматина, образованного спирализованной лишней Х-хромосомой. Тельце Барра обнаруживается при хромосомном наборе 46, XX (нормальные женщины); 47, ХХУ и 48, ХХУУ (классический синдром Клайнфельтера). Два тельца Барра обнаруживаются у человека, имеющего три Х-хромосомы, (47, XXX); три Х-хромосомы и одну У (48, ХХХУ, синдром Клайнфельтера); 49, ХХХУУ (синдром Клайнфельтера). Три тельца Барра встречаются при кариотипе 48, ХХХХ и 49, ХХХХУ (тяжелый синдром Клайнфельтера).
В полиплоидных клетках число телец полового хроматина соответствует плоидности. По формуле Гарднера, число телец Барра (В)
![]()
равно В = Х — , где Х — число Х-хромосом, Р — степень плоидности клетки. В неполиплоидных клетках число телец полового хроматина равно числу Х-хромосом минус единица (В = Х — 1).
Структурные изменения хромосом
Хромосомы могут подвергаться различным структурным изменениям. Особенно важное значение имеют потеря отдельных фрагментов хромосом (деления) или перенос участка одной хромосомы на другую (транслокация). Транслокация обозначается латинской буквой /, в скобках рядом с ней пишут индекс группы или номер хромосомы-донора, обозначение переносимого участка. Эти же обозначения указываются для хромосомы-реципиента, например 46, XXt (Ср + + В4q —). В скобках буквами р и q указывают плечи хромосом, затрагиваемые транслокацией. Короткое плечо хромосомы обозначают буквой р, длинное — буквой q, спутник — буквой s, и т. д. Увеличение длины плеча обозначается знаком плюс, а уменьшение — знаком минус (оба они ставятся после символа хромосомы).
Появление одной лишней хромосомы в кариотипе приводит к трисомии. Кратное увеличение числа всех хромосом носит название полиплоидии (могут быть триплоиды, тетраплоиды и т. д.). Потеря одной из пары гомологичных хромосом приводит к состоянию, которое называется моносомией. Изменения числа или строения хромосом называется хромосомными аберрациями.
Рассмотрим наиболее частые виды структурных нарушений хромосом — делеции и транслокации. При делеции общее количество хромосом не изменено. Однако в какой-то хромосоме недостает генетического материала, что вызывает различные изменения фенотипа. Чаще всего встречается делеция 5-й и 18-й аутосом и Х-хромосомы. Делеции приводят к развитию различных наследственных заболеваний и синдромов.
В 1963 г. Ж. Лежен описал синдром «кошачьего крика». Крик таких детей напоминает «мяуканье кошки». У детей резкое недоразвитие гортани, круглое лунообразное лицо, микроцефалия, микрогнатия, монголоидный разрез глаз, низко расположенные деформированные ушные раковины, мышечная гипотония, слабо выраженные вторичные половые признаки. Эти дети умственно отсталые. В кариотипе детей отмечается делеция короткого плеча 5-й пары хромосом.
Деления длинного и короткого плеча 18-й хромосомы сопровождается различными нарушениями строения лица, скелета, внутренних органов. У детей отмечается умственная отсталость, гипотрофия, гипотония, микроцефалия, недоразвитие лица, низкий грубый голос, недоразвитие наружных половых органов, среднего уха, атрезия наружного слухового прохода и другие аномалии.
При делеции короткого плеча 18-й хромосомы у больных также отмечаются различные дефекты со стороны скелета, внутренних органов и умственная отсталость.
Делеция короткого плеча Х-хромосомы может трактоваться как частичная моносомия по Х-хромосоме. Описана у женщин, у которых наблюдается задержка роста, недоразвитие яичников без тяжелых соматических аномалий. Хотя половой хроматин у них выявляется, однако его размеры значительно меньше, чем в норме.
При хронических миелолейкозах отмечается укорочение короткого плеча 21-й хромосомы (так называемая филадельфийская хромосома). Однако эта хромосома обнаруживается только в клетках крови и пунктате костного мозга. Другие же клетки имеют нормальный кариотип.
В результате двух концевых нехваток с последующим соединением разорванных концов образуются кольцевые хромосомы. Поэтому данное нарушение структуры хромосом фактически является частным случаем делеции. Клиническая картина больных — носителей кольцевых хромосом — напоминает таковую при делеции соответствующей хромосомы. Так, при кольцевой хромосоме группы В (5-я пара) развивается клиническая картина синдрома «кошачьего крика», а при кольцевой Х-хромосоме клиническая картина близка синдрому Шерешевского — Тернера.
Транслокации — это структурные перестройки, при которых происходит обмен генетического материала между хромосомами. Возможны различные виды транслокаций: реципрокные, при которых происходит взаимный обмен фрагментами; нереципрокные, когда генетический материал одной хромосомы переносится на другую, и наконец центрические соединения. Наиболее часто встречаются именно последние транслокации между акроцентрическими хромосомами. При этом утрачивается только небольшой фрагмент коротких плечей акроцентрических хромосом. Большую часть таких перестроек можно считать сбалансированной, так как они не вызывают серьезных отклонений в фенотипе носителя транслокации. Однако потомство таких носителей имеет клинически выраженные дефекты, характерные для аномального набора хромосом.
Известно, что болезнь Дауна может наблюдаться как при трисомии по 21-й аутосоме, так и при транслокации фрагмента этой хромосомы на другие. У таких больных хромосом 46, но одна из хромосом фактически двойная, так как к ней еще прикреплен фрагмент 21-й хромосомы и в результате такая перестройка оказывается не сбалансированной. У родителей этих больных кариотип включал 45 хромосом, но одна из хромосом была фактически двойной (с транслокацией). При оплодотворении яйцеклетки, содержащей эту хромосому, нормальным спермием в зиготе фактически будут три 21-х хромосомы, что фенотипически проявляется болезнью Дауна.
21-я хромосома чаще всего транслоцируется на 15-ю или на другие хромосомы группы Д (13-ю, 14-ю) у женщин, или на 22-ю у мужчин. В таком случае у молодых здоровых родителей может родиться ребенок с болезнью Дауна в отличие от трисомии 21-й хромосомы, которая чаще бывает у детей, рожденных пожилыми матерями. Определить наличие транслокации у индивидуума до рождения ребенка с болезнью Дауна без исследования кариотипа фактически невозможно, так как фенотип этих носителей мало чем отличается от фенотипов лиц с нормальными генотипами. Поэтому во всех этих случаях исследование кариотипа имеет особенно важное значение.
Механизм развития болезни Дауна при транслокации у одного из родителей можно представить следующим образом. При транслокации кариотип индивидуума состоит из 45 хромосом, так как одна хромосома увеличена в размере. Транслокация касается всех клеток, в том числе и оогоний и сперматогоний. При образовании половых клеток (гамет) в одну гамету попадает 23 хромосомы, а в другую 22. Но транслоцированная хромосома может оказаться как в гамете с 22 хромосомами, так и в гамете с 23 хромосомами. Таким образом, теоретически возможны 4 варианта гамет: 23 нормальные хромосомы, 23 с транслокацией, 22 нормальные хромосомы и 22 с транслокацией. Если транслокацию обозначить апострофом, то получится следующий ряд гамет: 23 231 22 221.
Если эти гаметы будут оплодотворены нормальной гаметой противоположного пола, то получим следующие комбинации: 1) 23 + 23 = = 46 хромосом (нормальный кариотип); 2) 231 + 23 = 461 хромосом, но фактически 47 хромосом (в данном случае разовьется болезнь Дауна); 3) 22 + 23 = 45 хромосом (такая зигота не жизнеспособна и погибает); 4) 221 +23 = 451 хромосом (в этом случае рождается индивидуум с транслокацией, как и один из его родителей).
Шансы родить ребенка с болезнью Дауна (при транслокации у одного из родителей) составляют 33%. Это очень большой риск и в таком случае дальнейшее деторождение не желательно, тем более что есть риск получить транслокацию и у внуков. Если рождается ребенок с болезнью Дауна, вызванной трисомией по 21-й хромосоме, у родителей с нормальным кариотипом, то шансы родить повторно такого же ребенка очень незначительны. Однако не во всех случаях при рождении ребенка с болезнью Дауна вследствие транслокации 21-й хромосомы транслокация имеется в соматических клетках матери. Примерно у половины матерей кариотип бывает нормальный, а транслокация произошла во время мейоза, предшествующего образованию яйцеклетки, из которой развился организм больного ребенка.
Глава 5. Мозаицизм.Это состояние, когда в организме перемешаны клетки с нормальным и аномальным кариотипами, скажем, 46/47 или 46/45. Возникает оно вследствие нерасхождения хромосом на начальных этапах эмбрионального развития. Мозаицизм дает стертые, слабо выраженные симптомы заболевания по сравнению с больными, у которых изменен кариотип во всех клетках. Больной с мозаичным вариантом болезни Дауна может иметь только некоторые физические признаки этого заболевания. Развитие интеллекта не нарушено. При мозиацизме 45ХО/46ХХ синдром Шерешевского — Тернера выражен более мягко. У таких больных возможно развитие тканей яичников и овуляция. При кариотипе 46ХУ/47ХХУ более мягко выражен синдром Клайнфельтера. Среди больных женщины- и мужчины-мозаики с указанными кариотипами встречаются чаще, чем «чистые» случаи синдрома Шерешевского — Тернера или синдрома Клайнфельтера. С возрастом клон аномальных клеток постепенно элиминируется, и поэтому трудно установить мозаицизм в пожилом возрасте, хотя в эмбриональном и раннем постэмбриональном периоде он был выражен достаточно и мог привести к развитию фенотипических признаков заболевания. Чем меньше в организме аномальных клеток, тем слабее выражены признаки заболевания. Этим можно объяснить стертые и рудиментарные формы данных заболеваний.[7]
При заболеваниях крови может происходить кратное (полиплоидия) или некратное (анэуплоидия) увеличение количества хромосом. Однако оно наблюдается только в клетках крови, а в других же соматических клетках кариотип нормальный.
Список использованной литературы.
1. Бердышев Г.Д., Криворучко И.Ф. Генетика человека с основами медицинской генетики. – Киев: Вища школа, 1979.
2. Бочков Н.П. Генетика человека. – Москва, 1973.
3. Фогель Ф. Мотульски А. Генетика человека: история хромосомы человека, формальная генетика. Москва: Мир, 1989.
4. Штерн, Курт Основы генетики человека. Москва: Медицина, 1965
5. Маккьюсик В. Генетика человека. Москва: Мир, 1967.
[1] Генетика человека с основами медицинской генетики. Бердышев Г.Д., Криворучко И.Ф. с.5-21
[2] Генетика человека: история хромосомы человека, формальная генетика. Фогель Ф. Мотульски А. с.11-19
[3] Генетика человека: история хромосомы человека, формальная генетика. Фогель Ф. Мотульски А. с.23-31
[4] Генетика человека. Бочков Н.П. с. 44
[5] Генетика человека. Маккьюсик. В. с.10, 13-22
[6] Основы генетики человека. Штерн, Курт. с.41-60
[7] Генетика человека. Бочков Н.П. с.90
Похожие работы
... индивидуальных хромосом; - накапливать сведения об изменениях хромосом в опухолевых клетках, у больных с наследственными заболеваниями крови и т.д. Главный недостаток методов, основанных на использовании низших организмов, заключается в невозможности экстраполировать полученные результаты на человека в связи с отсутствием процессов метаболической активации и детоксикации, характерных для всех ...



... к дочери. Голандрический признак передаётся от отца – сыну, т. е. Только по мужской линии. Геном - совокупность гаплоидного (1п) набора хромосом (23 хромосомы). 4. Мутационный процесс и наследственные заболевания человека: а) механизм генных мутаций. Болезни обмена веществ и молекулярные болезни человека. Наследование генных аномалий. Мутации происходят на каждом из перечисленных уровней, и ...
... человеческой линии эволюции с африканскими человекообразными обезьянами произошло значительно позже, чем 13 млн. лет назад. В последние годы антропогенез эффективно изучают также биомолекулярными методами. В основе этих методов изучения эволюции лежит допущение, что мера сходства двух таксонов соответствует мере их родства. Поэтому организмы, имевшие общего предка в недалеком прошлом, будут ...
... Далее она передавалась в рецессивной форме по материнской линии. Среди европейцев 0,01 % мужчин больны гемофилией. Генеалогическими методами установлена возможность наследования некоторых дарований человека, например способности к музыке, математике. Музыкальный талант в родословной Бахов проявлялся неоднократно. Степень проявления таланта, разумеется, зависит от сочетания других психофизических ...
0 комментариев