Марфенин Н. Н.
Накопленные к настоящему времени сведения о морфологии колониальных гидроидов ограничиваются описаниями отдельных “типичных” фрагментов колонии, таких как побег, гидрант, гонангий, столон. Такое положение можно было бы считать приемлемым, если бы колония не была целостным образованием. Однако подробные исследования интеграции колонии, выполненные в .последнее десятилетие на Dyпаmena pumila (L.), показали, что допущение о хаотичности строения колонии или представление колонии в виде суммы особей совершенно необоснованны (Марфенин, 1973, 1977; Карлсен, 1982). Более того, форма исследуемой части колонии, так же как и ее функционирование, в определенной степени зависит от места, занимаемого частью в колонии, и от состояния колонии в целом.
Таким образом, назрела необходимость в переописании морфологии колониальных гидроидов целой колонии, а не отдельных ее фрагментов. Описание целесообразно начинать с широко распространенных видов, таких как Obelia loveni. Этому и посвящена настоящая статья.
Исследование проводили на Беломорской биологической станции МГУ с июня но август 1980 г. Изучали морфологию только живых колоний. Для этого колонии выращивали из отдельных побегов на пластинах 8* 10 см (оргстекло), которые содержали как в лаборатории, так и в море — месте естественного обитания гидроида. Для этого пластины прикрепляли к буям, затопляемым во время прилива. В лаборатории колонии находились в кристаллизаторах с постоянной аэрацией. Воду меняли ежедневно. Температура свежей воды в момент смены равнялась лабораторной. Кормили свежевылупившимися науплиусами Artemia salina пропорционально (недозированно), в течение 2 ч ежедневно.
Для регистрации расположения основных компонентов колонии (побеги, столоны, верхушки роста, гидранты, гонозооиды) применяли модифицированный И. А. Косевичем метод картирования (Марфенин, 1980). Всего было изучено более 30 небольших колоний, включающих в сумме 428 побегов.
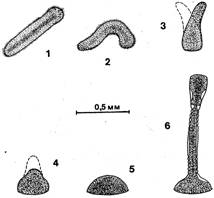
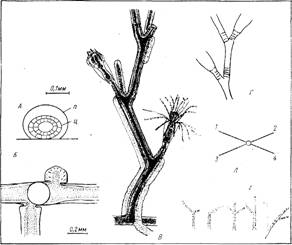
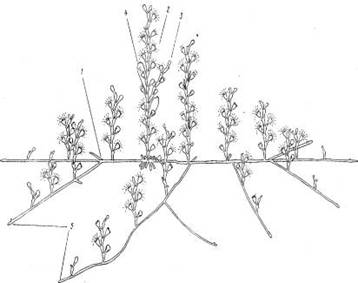
Место обитания и общий вид гидроида.О. loveni обитает преимущественно в зоне нижней литорали — верхней сублиторали на камнях, различных водорослях и искусственном субстрате. Колонии обелии, переплетаясь одна с другой, образуют густые заросли высотой около 2—4 см, выделяющиеся среди других гидроидов молочным цветом (при отсутствии обрастания) и особенно утонченной формой побегов (рис. 1).
Гидрориза.Нитевидная гидрориза представляет собой почти идеально цилиндрическую трубку 0,14—0,20 мм в сечении, стелющуюся по субстрату преимущественно прямолинейно. Перисарк гидроризы на всем протяжении гладкий, без зон кольчатости и без расширений под побегами (рис. 2,8). Ценосарк гидроризы редко заполняет все пространство внутри перисарка, занимая обычно лишь его центральную часть. Растущая верхушка столона всегда выпуклая на своем терминальном конце, имеющем простую полусферическую форму. Ценосарк в ней плотно прилегает к перисарку на протяжении 0,5—1,1 мм.
Боковые ветви гидроризы имеют такое же строение, как и материнский столон. Они отходят от последнего почти перпендикулярно, а при дальнейшем росте обычно слегка отклоняются к периферии колонии. Найти общие закономерности в порядке отхождения боковых столонов не удалось. Обычно ветвление происходит вблизи оснований побегов, хотя возможно их отхождение и в любом другом месте. Частота ветвления зависит в первую очередь от интенсивности питания. Анастомозы между соприкасающимися столонами образуются очень редко.
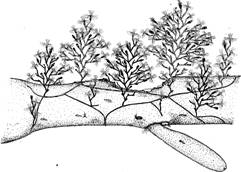
Рис. 1. Общий вид колонии О. loveni на аскофиллуме
Расстояния между побегами от 1,5 до 100 мм (в среднем 3 мм в естественных условиях и 3,5 — в лабораторных).
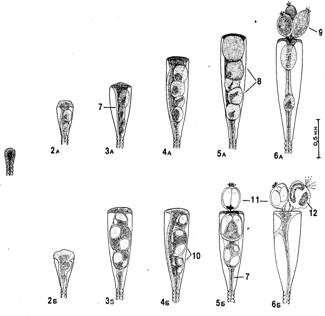
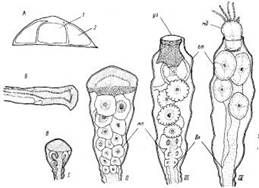
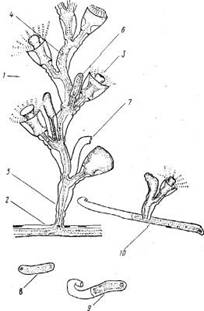
Побеги.В общем случае побег состоит из ствола и боковых ветвей, которые не различаются по строению (рис. 2). Высота ствола у побегов в исследованном нами материале обычно не превышала 30 мм, а число междоузлии, составляющих ствол, было до 25. Однако, по литературным сведениям, высота побегов у О. loveni может быть до 100 мм (Наумов, 1960); по нашим наблюдениям на Черном море, высота побегов О. loveni в обросте достигает 50 мм, а наиболее обычная (в мае) — 35—40 мм.
Суммарное число междоузлий побега, включая боковые ветви, может превышать у беломорских экземпляров О. loveni 200, т. е. почти в 10 раз больше, чем в стволе. Междоузлия устроены достаточно одинаково. По форме это почти идеально круглая в сечении трубка около 0,13 мм в диаметре, слегка изогнутая, длиной 0,6—0,8 мм. В междоузлии имеются проксимальная зона кольчатости (рис. 2, 10) из 1—8 (обычно 4—5) колец (2811 измерений), средняя гладкая зона и дистальная зона кольчатости (рис. 2, 9) из 3—4 колец, которая фактически является ножкой гидранта.
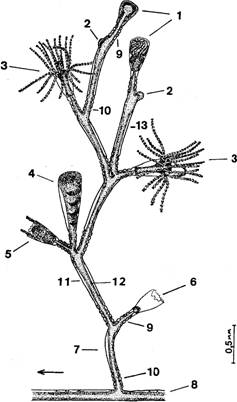
На границе гладкой зоны и дистальной зоны кольчатости (ножки гидранта) с выпуклой стороны под углом около 40° начинается следующее междоузлие, изогнутое в противоположную от предыдущего сторону. Радиус наибольшего изгиба междоузлия равен примерно 0,34 мм. Если не происходит постепенного смещения ориентации очередных междоузлий, образующихся по мере роста, то ствол побега оказывается билатерально симметричным. Плоскость симметрии перпендикулярна субстрату и параллельна столону, вернее столон лежит в этой плоскости.
Основание побега по своему строению ничем не отличается от основания междоузлия.
Рис. 2. Строение побега, зооидов и гидроризы: 1 — формирующиеся гидранты на разных стадиях развития, 2 — верхушки роста, 3 — гидранты с открытым и закрытым гипостомами, 4 — гонангий, 5 — сжавшийся гидрант с выставленными из гидротеки “сторожевыми” щупальцами, 6 — гидротека, оставшаяся после рассасывания гидранта, 7 — первое междоузлие всегда наклонено в дистальном направлении, 8 — столон (ветвь гидроризы), 9 — дистальная зона кольчатости междоузлия (ножка гидранта), 10 — проксимальпая зона кольчатости междоузлия, 11 — перисарк, 12 — ценосарк, 13 — боковая ветвь побега
У столона под побегом расши-рения не образуется. Первое междоузлие побега отходит от столона под углом примерно 70° и наклонено в своей базальной части к периферии колонии, но изогнуто так, что гидрант, венчающий это междоузлие, ориентирован к центру колонии (рис. 2,7).
Боковые ветви (ветви 1-го порядка) отходят от ножек гидрантов ствола с прилежащей стороны ножки (между стволом и гидрантом) (рис. 2, 13). Направление ветви немного не совпадает с плоскостью, проходящей через оси ствола и ножки гидранта. За счет этого “крона” побега не выглядит уплощенной. Первое междоузлие в ветви обычно изогнуто в сторону ствола. В строении и размерах ветвей и ствола различий не обнаружено. Часто одна из первых боковых ветвей растет значительно дольше остальных, становясь вторым стволом побега. Среди больших побегов такие раздвоенные встречаются как правило. Ветви 2-го порядка отходят от ветвей 1-го порядка так же, как последние от ствола. То же правило повторяется и у ветвей более высоких порядков. Встречаются побеги с ветвями 3, 4 и даже 5-го порядков.
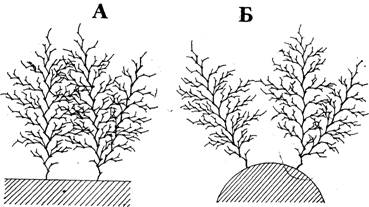
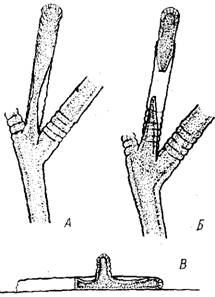
Рис. 3. Схема взаимного положения разветвленных побегов на плоском (А) и цилиндрическом (Б) субстрате. На плоском субстрате происходит переплетение ветвей побегов, на цилиндрическом — более эффективное использование окружающего пространства
При благоприятных условиях боковые ветви могут отходить почти от каждого междоузлия, кроме дистальных частей ствола и ветвей. Зарегистрированная нами частота ветвления, количественно определенная в виде отношения числа ветвей в побеге к суммарному числу междоузлии, колебалась от 0,06 до 0,39, явно не достигая при этом предела, так как питание колоний все же не было избыточным. Столь сильное разветвление, характерное для побегов О. loveni, приводит к тому, что у многих побегов более 80% междоузлий, а соответственно и гидрантов, и массы тела оказываются распределенными по ветвям и лишь около 20% сосредоточено в стволе.
В результате такого строения побег О. loveni обычно напоминает дерево с раскидистой кроной. Диаметр этой кроны достигает подчас 15 мм и более. Так как расстояние между основаниями побегов в среднем 3 мм, то большие побеги колонии, растущей на плоскости, должны переплетаться, увеличивая концентрацию гидрантов. Напротив, на цилиндрическом субстрате, таком как толстая веревка или таллом аскофилума, побеги должны в меньшей степени мешать друг другу, эффективно используя окружающее пространство (рис. 3).
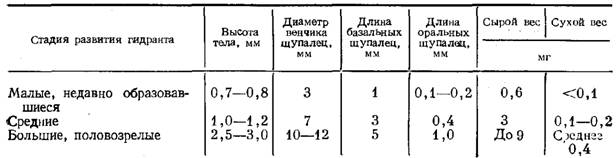
Гидранты.Голодные гидранты у О. loveni отличаются постоянством размеров (рис. 2,3). В расправленном состоянии тело гидранта около 0,5 мм длиной и 0,1—0,15 мм в диаметре. Булавовидный гипостом примерно 0,2 мм высотой и 0,15 мм диаметром в самой широкой части. 22—30 щупалец длиной в расправленном состоянии около 1 мм и 0,02 мм в поперечнике расположены своими основаниями в один ряд, хотя сами щупальца изогнуты так, что составляют как бы два яруса одного венчика — верхний и нижний. Последним гидрант опирается на гидротеку. Щупальца входят в верхний и нижний ярусы поочередно, ничем морфологически не различаясь.
Бокаловидная (или колоколовидная) гидротека бывает высотой от 0,4 до 0,65 мм и диаметром устья от 0,2 до 0,3 мм (средние размеры соответственно 0,5 и 0,28 мм). Соотношение высоты гидротеки и диаметра устья колеблется в пределах от 1,8 до 2,8. Эти размеры гидротеки достоверно отличаются от приведенных в литературе:
Д. В. Наумов (1960) сообщает, что длина гидротеки у О. loveni 0,6— 0,8 мм, а диаметр устья 0,3—0,37 мм. Это отчасти объясняется тем, что мы измеряли высоту гидротеки от устья до диафрагмы, так как граница между гидротекой и ножкой не всегда достаточно отчетливо видна.
Перисарк ножки гидранта четковидный. Длина ножки бывает от 0,25 до 0,7 мм. Ценосарк ножки занимает лишь центральную часть пространства внутри перисарка.
Гонангии.В период размножения, продолжающийся у беломорских О. loveni в течение всего лета, гонангии на колониях находятся на разных стадиях развития. Гонангии обычно имеются лишь на больших побегах. Они образуются чуть сбоку, на прилежащей к стволу стороне ножки гидранта, часто по два от одной ножки (рис. 2,4). Гонофоры (Гонофоры — медузоидные почки (Allman, 1871; Hyman, 1940) закладываются на одной стороне бластостиля. Более развитые расположены в дистальной части гонангия, менее развитые — в проксимальной.
Гонотеки мужских и женских гонангиев несколько различаются размерами. Высота мужской гонотеки 1,3—1,5, женской — 1,1—1,2 мм, диаметр устья примерно одинаков — 0,3—0,4 мм. Ножка четковидная, состоит из 3—4 колец. Мужской гонангий отличается от женского также большим числом гонофоров: у мужских их обычно 4—5 (до 10), а у женских обычно 3 и не более 6. Как правило, у женских медузоидов бывает много щупалец — более 10, тогда как у мужских их только 4. Содержимое мужских медузоидов однородное, а в женских хорошо видны яйца или формирующиеся планулы. По форме мужские медуозиды сливовидные, т. е. их диаметр меньше продольной оси, а женские тыквовидные (диаметр больше продольной оси).
Доля гонангиев в размножающейся колонии обычно бывает значительной. Так, например, в колонии с 21 побегом, на которых было 320 функционирующих и 73 развивающихся гидранта, оказалось 99 гонангиев, т. е. примерно один гонангии на 4 гидранта.
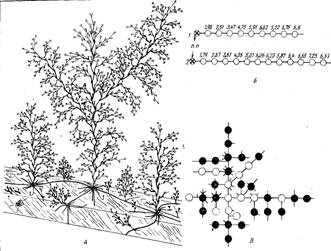
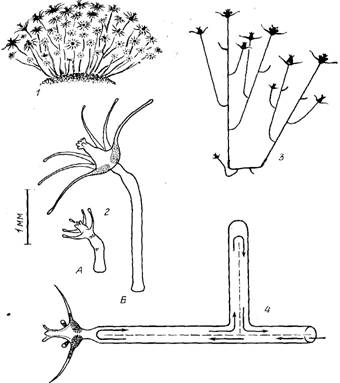
Строение колонии. Столоны и побеги располагаются в колонии в определенном порядке. От первичного побега отходят 3—4 главных столона (ветви гидроризы 1-го порядка). Они часто бывают наиболее длинными, если не упираются в какие-либо препятствия. От них отходят боковые столоны (ветви гидроризы 2-го порядка), длина которых обычно тем меньше, чем ближе к периферии колонии они находятся.
В расположении побегов по столону также наблюдается определенная закономерность. Так как новые побеги образуются только вблизи растущих верхушек столонов, то естественно, что самые старые (а следовательно, и самые большие) побеги находятся в центре колонии. Получается, что высота побегов на ветви гидроризы постепенно уменьшается к периферии (рис. 4). При сильном недостатке пищи старейшие побеги начинают рассасываться. В этом случае их размеры могут быть меньше, чем более молодых соседей. При нормальном питании рассасывание отдельных гидрантов и ветвей все равно в старых побегах более выражено, чем в молодых. У них постепенно рассасываются нижние гидранты и веточки, так что базальная часть ствола обнажается (рис. 4, 2).
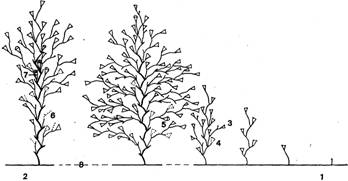
Рис. 4. Схема строения части колонии на одной ветви гидроризы: 1 — дистальная часть столона с формирующимися побегами, 2 — проксимальная часть столона с частично рассасывающимися побегами, 3 — функционирующие гидранты, 4 — рассосавшиеся гидранты, 5 — функционирующие гидранты второго поколения, 6 — рассосавшиеся части ветвей, 7 — гонангии, 8 — столон (пунктир здесь обозначает не показанные на рисунке части ветви гидроризы, в которых происходит постепенный переход от одного типа побега к другому)
Таким образом, часть колонии, включающая один столон с принадлежащими ему побегами, морфологически полярна. На одном ее конце находятся верхушка роста столона, молодые побеги и новообразовавшиеся боковые столоны; на другом — соединение с материнским столоном, старые побеги, гонангии на них и большие боковые столоны. По описанному строению радиальных частей колонии в целой колонии можно в общем случае условно выделить три зоны: центральную (или депрессии), где идет рассасывание побегов и, возможно, даже гидроризы; среднюю (или зону воспроизводства), в которой расположены побеги наибольших размеров (добывающие для колонии больше всего пищи) и происходит половое размножение; периферическую (или зону роста), где в наибольшей степени выражены рост колонии, освоение субстрата и образование побегов.
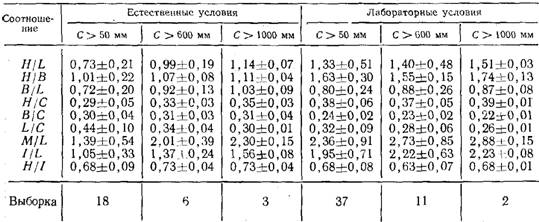
Количественные показатели структуры колонии. Строение целой колонии может быть также охарактеризовано с помощью соотношений основных ее показателей: общей длины ценосарка (С), длины гидроризы (L), числа функционирующих гидрантов (Н),числа верхушек роста (В). Отношения H/L, H/В, B/L, Н/С, В/С, L/C называются показателями структуры колонии (Марфенин, 1977), и каждое имеет свой функциональный смысл. Н/С дает представление о плотности расположения гидрантов в колонии; H/L — о проективной концентрации гидрантов на занятой поверхности субстрата; В/С — в степени разветвленности колонии; B/L — о проективной концентрации верхушек на занятой поверхности субстрата; Н/В косвенно свидетельствует об относительном количестве пищи, необходимом для обеспечения роста одной верхушки; L/C — о соотношении между побегами и гидроризой (косвенно — о высоте побегов). Вместе эти показатели дают достаточно полное представление об особенностях строения колоний разных видов гидроидов.
Показатели структуры колоний Obelia loveni (Allm.) (значения символов указаны в тексте)
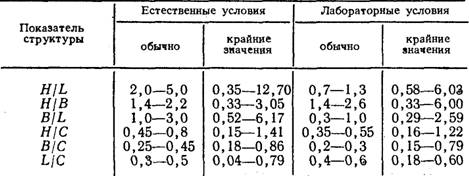
В наших опытах показатели структуры были рассчитаны на основании 86 картирований 19 колоний, культивируемых в естественных условиях, и 55 картирований 9 колоний, содержащихся в условиях лаборатории в течение двух месяцев — июля и августа. Результаты показали, что колонии в естественных и лабораторных условиях имели разное строение (таблица). Сравнение морфологии колоний двух серий показало, что они различаются степенью разветвленности побегов, их максимальной высотой, расстояниями между побегами и степенью развития гидроризы. Старые побеги в естественных условиях были более высокими и разветвленными, чем в лабораторных. Однако гидрориза была длиннее у лабораторных колоний, и расстояния между побегами здесь были достоверно больше.
В течение двух месяцев показатели структуры менялись без видимой закономерности, не выявляя месячной ритмики, обнаруженной ранее у Dynamena pumila (Марфенин, 1977).
Анализ морфологических показателей. Подробное изучение морфологии колонии дает возможность существенно расширить список морфологических показателей, используемых в систематике семейства Campanulariidae (Cornelius, 1982). В то же время в ряде случаев представляется возможным объяснить их функциональную роль в жизнедеятельности колонии. Это позволяет несколько приблизиться к оценке сравнительной значимости морфологических показателей, столь важной при моделировании формы любого организма. Мы полагаем, что колониальные гидроиды особенно перспективны как объекты моделирования из-за относительной простоты количественного описания их формы. Первая попытка подобного моделирования была предпринята сравнительно давно (Braverman. Schrandt, 1967) и выявила недостаток числа количественных показателей, используемых в описании колонии у гидроидов.
Мы надеемся, что приведенные ниже показатели окажутся полезными как в систематике гидроидов, так и при моделировании формы побега и колонии в целом.
1. Диаметр ствола побегов, ветвей, ножек гидрантов и столонов — основополагающая характеристика колонии, особенно в семействе Campanulariidae, так как вся колония фактически “построена из трубок”. Диаметр перисарка косвенно указывает на эффективность использования клеточного материала (чем больше диаметр, тем больше живых тканей в единице колонии), а также тесно связан с жесткостью конструкции. У О. loveni относительно тонкие стволы и гидрориза. Такие колонии гибки, но не обладают необходимой жесткостью для того, чтобы противостоять течению воды.
2. Переплетение ветвей побегов способствует усилению их общей жесткости. Переплетение достигается за счет значительного угла отхождения боковых ветвей от ствола и других ветвей, а также за счет интенсивного ветвления.
3. Прямолинейный рост гидроризы и ее ветвление под прямым углом способствуют относительно быстрому распространению по субстрату и освоению его.
4. Локализация мест ветвления гидроризы вблизи оснований побегов приводит к увеличению прочности прикрепления колоний к субстрату, что особенно важно при увеличении размеров и соответственно парусности побегов. Специальных расширений столонов под побегами, свойственных, например, семейству Sertulariidae, у Obelia не выработалось. Такие расширения столонов связаны с закреплением мест ветвления гидроризы исключительно под побегами.
5. Отсутствие анастомозов в гидроризе тесно скоррелировано с умеренностью ее ветвления, а это означает преобладание стратегии прямолинейного роста столонов и захвата новых площадей субстрата перед стратегией сдерживания экстенсивного роста и наиболее полным использованием занятой территории. Последнее присуще таким гидроидам, как Clava и Hydractinia, обитание которых обычно приурочено, по нашим наблюдениям, к определенным местам субстрата.
6. Доля междоузлиq и гидрантов, приходящихся на боковые ветви побега, демонстрирует значение ветвления, которое способствует концентрации гидрантов, т. е. увеличению их числа на единицу площади, занимаемой колонией. Доля гидрантов в стволе и боковых ветвях разного порядка показывает роль разветвления колонии и характер распределения функционирующих гидрантов в кроне, которое может быть равномерным или, к примеру, в основном периферическим — по поверхности кроны.
7. Высота побега, ширина кроны, длина междоузлия, число порядков разветвления, угол отхождения боковых ветвей от ствола, угол наклона боковых ветвей к плоскости “гидрант — ствол”, частота ветвления, продолжительность роста боковых ветвей — основные количественные показатели, с помощью которых можно точно моделировать форму побега.
8. Расстояния между побегами, частота ветвления гидроризы, место и угол отхождения боковых столонов от материнского — показатели, необходимые для моделирования колонии в целом.
9. Продолжительность жизненного цикла гидранта и число возможных жизненных циклов на одном междоузлии — функциональные показатели, необходимые для моделирования колонии в развитии.
Анализ тотальной морфологии колонии. В систематике гидроидов до сих пор используются почти исключительно показатели морфологии отдельных “типичных” фрагментов, как правило зооидов. Морфология побега описывается весьма приблизительно почти без использования количественных показателей. Естественно, что при таком подходе различия между побегами не заметны, а колония выглядит как простая сумма “одинаковых” побегов с одинаковыми зооидами.
Более подробное изучение целых колоний с применением методики их количественного картирования позволяет выявить различия между побегами и обнаружить закономерности в строении колонии. Различия между побегами выражаются в числе междоузлии и числе гидрантов, в разветвленности, а следовательно, и в числе верхушек роста, в наличии и числе гонозооидов.
Строение побегов меняется в пределах ветви гидроризы, на которой они находятся, постепенно и закономерно от одного ее конца к противоположному, что создает эффект морфологической полярности, или градиента, впервые отмеченный А. Г. Карлсеном (1982) на Dynamena pumila.
Ветви гидроризы с побегами у О. loveni соединены между собой определенным образом, так, что общее строение колонии можно назвать “центробежным”. Морфологическая полярность ветвей гидроризы вследствие “центробежного” строения колонии отражается и в ее общем строении. В колонии можно различить три зоны: центральную (или зону депрессии) и расположенные концентрически вокруг нее среднюю зону (или зону воспроизводства) и периферическую зону (роста).Часто зона депрессии еще отсутствует, и тогда можно выделить только две зоны — периферическую и зону воспроизводства, которая в этом случае занимает центральное положение. Подобное подразделение колонии на две зоны было предложено нами ранее для Dynamena pumila (Марфенин, 1973). Теперь мы убедились, что это лишь частный вариант, а более общий — дифференциация колонии на описанные выше три морфофункциональные зоны.
Таким образом, колония в целом имеет определенное, довольно закономерное строение, для описания которого удобно использовать количественные показатели.
Список литературыКарлсен А. Г. 1982. Саркон как уровень организации колониальных гидроидов (Leptolida, Hydrozoa). — Докл. АН СССР, 263, № 2, 510—512. Марфенин Н. Н. 1973. Морфология роста в колонии гидроидного полипа Dynamena pumila (Hydrozoa. Leptolida). — Журн. общ. биол., 34,№ 5, 727—737. М а р ф е нин Н. Н. 1977. Опыт изучения интеграции колонии гидроида Dynamena pumila (L.) с помощью количественных морфологических показателей. — Журн. общ. биол., 38, № 3, 409—422. Марфенин Н. Н. 1980. Метод картирования пространственной организации колониальных Hydrozoa и его значение при изучении частей колонии.— В кн.: Теоретическое и практическое значение кишечнополостных. Л., с. 66—69. Наумов Д. В. 1960. Гидроиды и гидромедузы морских, солоноватоводных и пресноводных бассейнов СССР. М.—Л. Allman G. J. 1871. A monograph of the Gymnoblastic or Tubularian Hydroids. I. The hydroida in general. L. Cornelius P. F. S. 1982. Hydroids and hydromedusae of the family Campanulariidae recorded from the eastern North Atlantic, with a world synopsis of genera. — Bull. Brit. Mus. nat. Hist. (Zool.), 42 (2), 37—148. Braverman M., Schrandt R. 1967. Colony development as a problem in pattern formation. — Gen. Syst, 12, 39—51. Ну man L. 1940. The Invertebrates: I. Protozoa through Ctenophora. N. Y., p. 400
Похожие работы
... использовали. Этот процесс был изучен довольно подробно у Campanularia (Obelia) flexuosa (Crowell, 1953). Наше исследование позволяет уточнить сведения о жизненном цикле гидрантов применительно к Obelia loveni. В жизненном цикле гидранта целесообразно различать четыре этапа: 1) формирование, 2) функционирование, 3) рассасывание и 4) диапауза. 1. Формирование гидранта от момента появления почки ( ...
... и процессов интеграции колоний у гидроидов при лабораторной культивации на примере Dynamena puinila (L.) //Теоретическое и практическое значение кишсчнополостных. Л. С. 34—40. Косевнч И. А. 1984. Диморфизм побегов в колонии гидроида Obelia longissima (Pallas, 1766) (Hydrozoa, Thecaphora, CampanuJariidac). Деп. в ВИНИТИ 16.03.84 1984, № 1507. Марфенин Н. Н. 1974. Структура колонии гидроидного ...
... колоний Cordylophora caspia Pall. В результате гидранты оказываются расположенными не по поверхности кустовидного побега, а равномерно внутри его, что больше напоминает моноподиальные колонии с терминально расположенными зонами роста или симподиальные колонии. Можно привести и еще примеры, доказывающие, что ,в пределах данного типа колоний возможны весьма широкие вариации в строении. Однако общей ...
... к гидродинамическим условиям существования. На эти вопросы пока нет исчерпывающих ответов. Однако подробное изучение морфологии колоний Obelia loveni [8], О. longissima [3] и О. geniculata дает возможность высказать ряд предположений о функциональной подоплеке различных особенностей строения гидроидов. Тем самым мы стремимся конкретизировать проблему, указать на возможные пути ее решения и ...
0 комментариев