ПЛАН
Введение
Новые варианты вируса СПИД
Статистика
Строение вирусной частицы ВИЧ
Строение вирусного генома и экспрессия генов ВИЧ
Гены и белки ВИЧ
Регуляция экспрессии вирусных генов
Теории происхождения ВИЧ
Передача ВИЧ-инфекции
Ко-факторы ВИЧ-инфекции
Патогенез и клиника ВИЧ-инфекции
Патогенез
Механизмы взаимодействия ВИЧ с различными звеньями иммунной системы
Влияние ВИЧ на Т-хелперы
Взаимодействие ВИЧ с Т-супрессорами
Результаты взаимодействия ВИЧ с В-клетками
Макрофаги и ВИЧ
Взаимодействие ВИЧ с моноцитами
Взаимодействие ВИЧ с другими клетками организма
Система интерферона при ВИЧ-инфекции
Клинические проявления
Персистирующая генерализованная лимфаденопатия
СПИД-ассоциированный комплекс
Оппортунистические инфекции и опухоли
Особенности инфицирования и заболевания детей
Вирус иммунодефицита человека типа 2 (ВИЧ 2)
Географическое распространение
Строение вируса
Эпидемиология
Клинические отличия
Лабораторная диагностика
Диагностика ВИЧ-инфекции
Лечение
Возможности разработки вакцин
Заключение
ВВЕДЕНИЕ
ВИЧ-инфекция, подобно пожару, охватила сейчас почти все континенты. За необычайно короткое время она стала проблемой номер один для Всемирной организации здравоохранения и ООН, оттеснив на второе место рак и сердечнососудистые заболевания.
Пожалуй, ни одна болезнь не задавала ученым такие серьезные загадки за столь незначительный срок. Война с вирусом СПИДа ведется на планете с нарастающими усилиями. Ежемесячно в мировой научной прессе публикуются новые сведения о ВИЧ-инфекции и ее возбудителе, которые часто заставляют коренным образом менять точку зрения на патологию этого заболевания.
Пока загадок больше...
Прежде всего - неожиданность появления и быстрота распространения ВИЧ. До сих пор не решен вопрос о причинах его возникновения. До сих пор неизвестна средняя и максимальная продолжительность его скрытого периода.
Установлено, что имеется несколько разновидностей возбудителя СПИДа. Изменчивость его уникальна, поэтому есть все основания ожидать, что обнаружатся очередные варианты возбудителя в разных регионах мира, а это может резко осложнить диагностику.
Еще загадки: какова связь СПИДа у человека со СПИД-подобными заболеваниями у животных (обезьян, кошек, овец, крупного рогатого скота) и какова возможность встраивания генов возбудителя СПИДа в наследственный аппарат зародышевых клеток?
Далее. Правомерно ли само название? СПИД расшифровывается как синдром приобретенного иммунодефицита. Иными словами, главный признак болезни - поражение иммунной системы. Но каждым годом накапливается все больше данных, доказывающих, что возбудитель СПИДа поражает не только иммунную, но и нервную систему.
С совершенно непредвиденными трудностями сталкиваются при разработке вакцины против вируса СПИДа.
К особенностям СПИДа относится то, что это, по-видимому, первый в истории медицины приобретенный иммунодефицит, связанный с конкретным возбудителем и характеризующийся эпидемическим распространением. Вторая его особенность - почти “прицельное” поражение Т-хелперов. Третья особенность - это первое эпидемическое заболевание человека, вызванное ретровирусами. В-четвертых, СПИД по клиническим и лабораторным особенностям не похож ни на какие другие приобретенные иммунодефициты.
ОПРЕДЕЛЕНИЕ ПОНЯТИЯ СПИД.
Термин СПИД впервые появился в Еженедельном Отчете о Заболеваемости и Смертности Центров по Контролю Заболеваний в 1982 году чтобы описать “...заболевание, сдержанно предполагающее о дефекте клеточного звена иммунитета, встречающееся без известных причин для уменьшения резистентности к той болезни...”
Начальный список Центра Контроля Заболеваний о СПИД-определяющих условиях, который включал саркому Капоши, пневмоцистоз, комплекс Mycobacterium avium и другие условия, несколько раз был модифицирован со значительными изменениями (Центр Контроля Заболеваний (ЦКЗ), 1985а, 1987а, 1992а).
В настоящее время ЦКЗ определяет СПИД во взрослом и подростковом возрасте 13 лет и старше как присутствие одного из 25 СПИД-индикаторных условий, таких как саркома Капоши, пневмоцистоз или диссеминированный комплекс Mycobacterium avium. У детей младше 13 лет определение СПИДа сходно с таковым у подростков и взрослых, за исключением того, что в список СПИД-определяющих условий входят также лимфоидный пневмонит и хронические бактериальные инфекции (ЦКЗ, 1987b). Далее будет приведен список ВИЧ-маркерных заболеваний. Область определения среди взрослых и подростков была расширена в 1993 году включением в этот список ВИЧ-инфекции у человека со снижением в крови количества CD4+ T-лимфоцитов менее 200 в 1 мм3. Текущие наблюдения заменили критерии, опубликованные в 1987 году, которые были основаны на клинических симптомах, а не на определении CD4+ T-лимфоцитов.
Итак, термин СПИД расшифровывается как синдром приобретенного иммунодефицита, но приобретенных иммунодефицитов много, а СПИД - один. Поэтому сегодня правильнее сказать так: СПИД - это такой приобретенный иммунодефицит, который отличается от других наличием определенного комплекса свойств и специфического возбудителя. Об этом специфическом возбудителе далее и пойдет речь.
ИСТОРИЯ ОТКРЫТИЯ ВИЧ.
Зимой 1980-81 года в госпиталь Нью-Йоркского университета поступили несколько человек с незнакомой для врачей формой саркомы Капоши - заболевания, открытого еще в 1872 году Моритцем Капоши. На коже нижних конечностей появляются узелки коричневато-красного или голубовато-красного цвета. Иногда они изъязвляются и омертвевают, но, обычно не поражают внутренние органы, и не считаются злокачественными опухолями (у большинства больных саркома Капоши длится 8-1 лет и хорошо поддается химиотерапевтическому лечению).
В США и странах Западной Европы саркома Капоши наблюдается чрезвычайно редко: 1-2 случая на 10 миллионов населения, причем, как правило, только у мужчин старше 60 лет. Мужчины же поступившие в Нью-Йоркский госпиталь были в возрасте 30 лет. Все они оказались гомосексуалистами. Саркома Капоши протекала у них злокачественно и большая часть из них погибла в течение 20 месяцев.
Весной 1981 года врачи Лос-Анджелеса обнаружили еще одну категорию больных - со злокачественной формой пневмоцистной пневмонии. Это заболевание вызывается простейшим Pneumocystis carinii и встречается крайне редко у лиц с подавленной функцией иммунной системы - например, подвергшихся интенсивной иммунодепрессивной терапии после трансплантации органов. Пневмоцистная пневмония была зарегистрирована у молодых людей, которые тоже оказались гомосексуалистами.
Летом 1981 года в США насчитывалось уже 116 подобных случаев...
Хотя клиническая картина указывала на известный уже к тому времени синдром иммунодефицита, причина и пути заболевания оставались неясными. Неожиданное появление болезни, молниеносное распространение, странная связь со злокачественными опухолями, пневмоцистной пневмонией, гемофилией, гомосексуализмом, венерическими болезнями, необычайно длительный скрытый период и отсутствие эффективных средств лечения - все это вызвало шок у врачей и ученых. Вирус, известный ныне как возбудитель СПИДа, был открыт только в 1983 году, и его называли по-разному.
Группа ученых Национального института рака в США, руководимых известным иммунологом и вирусологом Робертом Галло, открыла возбудителя Т-клеточного лейкоза, заболевания, зарегистрированного в конце 70-х годов в странах Карибского бассейна и в Южной Японии. Лейкоз протекал очень тяжело: больные погибали за 3-4 месяца.
Успеху Галло способствовало то, что в середине 70-х годов он обнаружил фактор роста Т-клеток, который сейчас называют интерлейкином-2. Это позволило культивировать Т-лимфоциты в пробирке.
Возбудителем острого Т-клеточного лейкоза у человека оказался ретровирус. Галло назвал “свой” агент вирусом Т-клеточной лейкемии человека - HTLV-1. И предположил, что он возник в Африке, где им заразились приматы Старого Света, да и человек тоже, что в Америку и страны Карибского бассейна вирус проник благодаря работорговле. Выяснилось, что у многих видов африканских обезьян в крови содержатся антитела к HTLV-1. Некоторые разновидности вируса, особенно выделенные у зеленых мартышек и шимпанзе, имели много сходного с HTLV-1. В дальнейшем оказалось, что HTLV-1 передается при переливании крови. Выделен еще один вирус этой группы, вызывающий редкое заболевание крови, - HTLV-2.
Как раз в это время в США началась эпидемия СПИДа, одним из путей передачи которого также было переливание крови. Поэтому Галло предположил, что HTLV-1 - возбудитель СПИДа. И действительно у некоторых больных удалось выделить антитела к HTLV-1, а у части удалось выделить и сам вирус. Однако Галло ошибся.
Группа ученых Пастеровского института в Париже под руководством Люка Монтанье, оснащенная значительно хуже группы Галло, была создана с целью изучить возможную связь ретровирусов с опухолевыми заболеваниями иммунной системы, выражающимися в увеличении лимфатических узлов (лимфаденопатии). Для выявления искомых вирусов французские ученые использовали открытый группой Галло интерлейкин-2. При исследовании одного пациента, болевшего лимфаденопатией в течение нескольких лет, был выделен вирус, идентифицированный как ретровирус. По своим свойствам он был похож на HTLV-1, но имел некоторые особенности. Затем аналогичный вирус выделили от больных СПИДом.
В 1983 году журнал “Сайенс” напечатал статью французских ученых. Они сообщали о наличии у 2 из 33 больных СПИДом ретровируса, который, в отличие от HTLV-1, не обладал способностью влиять на злокачественное перерождение Т-лимфоцитов. Авторы дали ему название LAV (вирус, ассоциированный с лимфоаденопатией). Он вызывает не размножение, а, наоборот, гибель Т-лимфоцитов.
Между обеими группами ученых начался интенсивный обмен идеями, биологическими материалами. Группа Галло, пользуясь разработанными ею методами, выделила от больных СПИДом новый ретровирус, названный HTLV-3. Удалось получить особую линию Т-лимфоцитов, в которой вирус интенсивно размножался, но Т-клетки не погибали. В начале 1984 года американцы сообщили об открытом ими вирусе в печати. И тогда же установили, что HTLV-3 и LAV идентичны. Поэтому вирус стали обозначать как HTLV-3/LAV. В 1986 году Комитет по таксономии и номенклатуре вирусов предложил дать возбудителю СПИДа новое название - HIV/ВИЧ (вирус иммунодефицита человека).
Итак, возбудитель СПИДа был открыт через два года после опубликования первых статей о новом заболевании, а метод его диагностики разработан менее чем через 2.5 года после первых публикаций. Это привело к тому, что на первой Международной конференции по СПИДу, состоявшейся в апреле 1985 года в Атланте господствовала атмосфера оптимизма.
Однако новые тревоги возникли при изучении уникальных особенностей возбудителя.
ОСОБЕННОСТИ ВОЗБУДИТЕЛЯ СПИДа.
ВИЧ был открыт. Роберт Галло и его сотрудники сначала полагали, что HIV и HTLV-1 принадлежат к одному семейству - HTLV, так как у них много общих свойств. Оба возбудителя обладают сродством к Т-лимфоцитам-хелперам с молекулой-рецептором CD4 на поверхности. Оба вируса переносятся при переливании крови. В обоих случаях вирусоносительство сопровождается накоплением соответствующих антител в крови. Есть сходство в строении оболочечных (env) генов.
Однако по мере изучения ВИЧ выяснились и его существенные отличия от HTLV-1. Они оказывают диаметрально противоположное патологическое действие: HTLV-1 превращает нормальную Т-клетку в злокачественную и вызывает безудержное размножение Т-хелперов, тогда как ВИЧ убивает эти клетки. Неодинаковым оказалось и их строение. Исследования А.Ф.Быковского и Л.Монтанье показали, что у сердцевины HTLV-1 сферическая форма, а у ВИЧ она конусовидная.
Существенные различия оказались и в геномах вирусов. По сравнению с HTLV-1 геном ВИЧ содержит несколько дополнительных генов. Главный белок сердцевины ВИЧ - р24 - не имеет аналогов у других ретровирусов. Процесс транскрипции генома ВИЧ протекает в тысячу раз быстрее, чем у клеточных генов, что в значительной степени объясняет поразительную скорость размножения ВИЧ.
Установлено, что ВИЧ стоит ближе всего к лентивирусам, вызывающим тяжелые хронические инфекции у копытных животных. Особенно отчетливо видны сходства ВИЧ с вирусом мэди-висна, который дает хроническую инфекцию у овец, приводящую, как и СПИД у людей, к смертельному исходу.
Подобно мэди-висна, ВИЧ характеризуется крайней изменчивостью - она в 30 -100, а по некоторым данным и в миллион раз выше, чем у вируса гриппа. Касается она не только штаммов вируса выделенных от разных больных, но и в разное время года от одного и того же больного. Это свойство резко затрудняет возможность получения вакцин против ВИЧ.
НОВЫЕ ВАРИАНТЫ ВИРУСА СПИД.
В апреле 1986 года в журнале “Сайенс” было опубликовано сообщение, что группа американских ученых во главе с Эссексом, выделила у здоровых людей в Сенегале вирус, сходный с вирусом африканских зеленых мартышек: сыворотки зараженных людей реагировали с антигенами вируса мартышек, но не давали реакции (или они были слабыми) с антигенами ВИЧ. Предполагалось, что вирус мог быть предшественником ВИЧ. Авторы наименовали выделенный вирус HTLV-4.
HTLV-4, подобно ВИЧ, инфицирует Т-хелперы, но не убивает их. Электронный микроскоп не зафиксировал его отличий от ВИЧ. По мнению Эссекса, хотя HTLV-4 инфицирует Т-хелперы у клинически здоровых людей, не исключено, что в последующем они заболеют.
В конце 1986 года Монтанье и сотрудники объявили об открытии нового вируса у двух больных СПИДом. По своему строению он не отличался от ВИЧ, тоже убивал Т-хелперы. Но, в противоположность ВИЧ, в сыворотках больных отсутствовали антитела к последнему и ДНК обоих вирусов не были идентичны. Авторы обозначили новый вирус как HIV-2 (ВИЧ-2). Сыворотки больных при ВИЧ-2 реагировали с вирусом зеленых мартышек, поэтому Монтанье считает, что у них может быть общее происхождение.
Сравнительное изучение геномов ВИЧ-1 и ВИЧ-2 показало, что в эволюционном плане ВИЧ-2 далеко отстоит от ВИЧ-1. Авторы высказывают предположение, что оба вируса существовали задолго до возникновения современной эпидемии СПИДа. В некоторых странах Западной Африки встречены типичные случая СПИДа при отсутствии антител к ВИЧ. Больных выделены ретровирусы, которые подобно ВИЧ-1, обладают сродством к CD4-хелперам и вызывают их гибель. ВИЧ-2 обнаруживается главным образом в Западной Африке. ВИЧ-2 родственен возбудителю СПИД-подобного заболевания у макак. Он способен инфицировать разные виды приматов, удаленные от человека на лестнице эволюции, тогда как ВИЧ-1 заражает только людей и шимпанзе. Подробнее о ВИЧ-2 будет сказано ниже.
Есть мнение, что непатогенный вирус HTLV-4 эволюционировал в безвредный вирус африканских зеленых мартышек, который превратился в патогенный ВИЧ-2 - предшественник ВИЧ-1.
Изучение СПИД стимулировало поиски сходных возбудителей. Помимо ВИЧ-1, в разных странах, преимущественно в Западной Африке, “обитают” различные ВИЧ-подобные варианты вируса, патогенные и непатогенные для человека. С другой стороны в этих же странах циркулируют варианты обезьяньих вирусов - патогенные для обезьян и не патогенные для них. Возможно, что новые ВИЧ-подобные вирусы человека больше сходны по своим биологическим и антигенным свойствам с обезьяньими вирусами, чем с ВИЧ.
Можно предположить, что по мере исследований будут найдены неизвестные ранее разновидности ВИЧ-подобных вирусов человека...
Сообщение об одном из таких открытий появилось в 1990 году. Предполагается, что выделен новый тип вируса иммунодефицита человека - ВИЧ-3.
Вирус был выделен от клинически здоровой женщины, которая была половым партнером положительного по антителам к ВИЧ мужчины. Сыворотка женщины была слабоположительной в ИФА, с низким титром в непрямом иммунофлюоресцентном тесте с антителами к ВИЧ-1 и давала слабые полосы в положениях, характерных для белков р24 и gp41 в иммуноблоте. Анализ выделенного ретровируса с помощью модифицированного теста на связывание антигенов показал, что выделенный изолят не является ВИЧ-1. Сравнение белков выделенного изолята - ANT70 - с белками ВИЧ-1 и ВИЧ-2 показало, что их молекулярная масса отличается от соответствующих белков ВИЧ-1 и ВИЧ-2. Сыворотки женщины, а потом и ее партнера лучше реагировали с ANT70, чем с ВИЧ-1 и ВИЧ-2. Позже вирус ANT70 был выделен и от полового партнера женщины.
Нуклеотидная последовательность генома выделенного вируса существенно отличается от последовательностей геномов ВИЧ-1 и ВИЧ-2. Последовательность длинного концевого повтора (LTR - long terminal repeat) на 3` конце вирусного генома отличается примерно на 30% от оснований LTR ВИЧ-1 и более чем на 50 % от LTR ВИЧ-2. Если учесть, что у разных штаммов ВИЧ-1 LTR различаются примерно на 15%, то обнаруженные у выделенного штамма отличия в 30% и более дают основания полагать, что выделен новый, третий представитель семейства вирусов-возбудителей СПИД - ВИЧ-3.
СТАТИСТИКА.
По данным ВОЗ на конец 1994 года в мире зарегистрировано 17 миллионов ВИЧ-инфицированных. Причем 66% из них находится в Африке на территориях южнее Сахары (11.2 миллиона человек). В Южной и Юго-Восточной Азии насчитывается около 3 миллионов носителей вируса СПИД. Во всей Австралазии зарегистрировано только около 12.000 зараженных. В 15 странах (все они расположены на территориях южнее Сахары) количество ВИЧ-инфицированных составляет в районе 500 человек на 10.000 населения. В 50 странах этот показатель колеблется в районе 5 человек на 10.000 населения. В остальных странах он ниже. Таким образом, наблюдается очень неравномерное распространение вируса СПИД, но все же заболевание имеет масштабы пандемии.
Различается и частота выявления вируса среди разных слоев населения. Среди мужчин-гомосексуалистов она составляет 60-90%, а среди наркоманов, употребляющих наркотики внутривенно - 13-20%. Хотя в последнее время начинает наблюдаться обратная картина. Первая волна эпидемии распространялась среди мужчин-гомосексуалистов, а нынешняя - среди наркоманов, применяющих внутривенные инъекции, что позволит эпидемии в большей мере захватить и гетеросексуальную часть населения. Переход эпидемии на гетеросексуальную часть населения будет происходить благодаря бисексуальным мужчинам, наркоманам и проституткам. Сегодня процент женщин среди заболевших СПИДом составляет около 5-10%, среди которых 50% заболевших - инъекционные наркоманы, 29% заразились при гетеросексуальных половых контактах. Правда, изучение гетеросексуального пути распространения выявило различия в эффективности передачи вируса - она максимальна среди женщин, партнеры которых больны СПИДом, а для передачи от больной женщины к ее партнерам составляет 65%.
Накоплены также убедительные данные о том, что в Африке ВИЧ распространяется в основном благодаря гетеросексуальным половым контактам, причем соотношение между числом заболевших мужчин и женщин составляет примерно 1:1. Помимо половых контактов важную роль играют переливания зараженной крови, и, возможно, иглы для лечебных манипуляций, а также вертикальный путь передачи. Сейчас установлено, что первые случаи СПИДа имели место в Африке еще в конце 70-х годов. Эпидемиологические данные для ряда африканских стран показали, что в определенных группах процент зараженных очень высок: 80-90% проституток, 30% их клиентов, 30% больных посещающих венерологические отделения, 10% доноров крови, 10% женщин, посещавших клиники пренатального профиля. И хотя высокий уровень инфицированности был вначале характерен только для районов Центральной Африки, вирус и вызываемая им болезнь распространились оттуда почти по всему континенту.
Далее приводятся некоторые конкретные цифры и показатели по распространенности ВИЧ/СПИД на нашей планете.
Число зарегистрированных ВОЗ случаев СПИДа на разных континентах
Основано на сообщении от 15 декабря 1995 года.
НОВЫЕ СЛУЧАИ СПИДа
ГОД Африка Америка Азия Европа Океания ВСЕГО
1979 0 2 0 0 0 2
1980 0 185 1 17 0 203
1981 0 322 1 20 0 343
1982 2 1156 1 80 91 1330
1983 17 3352 8 295 6 3678
1984 187 6680 8 570 76 7521
1985 521 12682 27 1475 142 14847
1986 5438 21322 86 2395 252 29493
1987 16854 34562 150 9640 324 61530
1988 28212 47697 176 10811 598 87494
1989 41295 56202 288 14355 699 112839
1990 54528 65041 478 17311 770 138128
1991 72756 78579 838 18937 897 172007
1992 73631 99881 2039 20697 866 197114
1993 67124 100731 7368 22053 879 198155
1994 65684 83475 11707 23541 906 185313
1995 16486 47793 5454 11906 174 81813
________________________________________________________
ВСЕГО 442735 659662 28630 154103 6680 1291810
ВСЕГО СЛУЧАЕВ СПИДа
ГОД Африка Америка Азия Европа Океания ВСЕГО
1979 0 2 0 0 0 2
1980 0 187 1 17 0 205
1981 0 509 2 37 0 548
1982 2 1665 3 117 91 1878
1983 19 5017 11 412 97 5556
1984 206 11697 19 982 173 13077
1985 727 24379 46 2457 315 27924
1986 6165 45701 132 4852 567 57417
1987 23019 80263 282 14492 891 118947
1988 51231 127960 458 25303 1489 206441
1989 92526 184162 746 39658 2188 319280
1990 147054 249203 1224 56969 2958 457408
1991 219810 327782 2062 75906 3855 629415
1992 293441 427663 4101 96603 4721 826529
1993 360565 528394 11469 118656 5600 1024684
1994 426249 611869 23176 142197 6506 1209997
1995 442735 659662 28630 154103 6680 1291810
______________________________________________________
ВСЕГО 442735 659662 28630 154103 6680 1291810
Данные отностительно конкретных стран можно найти в приложении.
Но, к сожалению, точные цифры числа заболевших и инфицированных в мире неизвестны. Это обусловлено несколькими причинами. Во-первых, сама статистика несовершенна - ВОЗ регистрирует только больных с выраженной картиной заболевания и не учитывает лиц с пре-СПИДом и вирусоносителей. Во-вторых, некоторые страны дают неполные данные потому, что у них тестированию на ВИЧ подвергается незначительная часть людей, относящихся к категории высокого риска. Это в основном страны Африки и Азии, где отсутствие средств для постановки соответствующих исследований препятствует полноценному выявлению инфицированных, прежде всего доноров крови. Третья причина состоит в том, что правительства ряда стран опасаются публикации этих сведений, чтобы они не навредили иностранному туризму, который является там одним из главных источников национального дохода.
СТРОЕНИЕ ВИРУСНОЙ ЧАСТИЦЫ ВИЧ.
Вирион имеет сферическую форму, диаметром 100-150 нм. Основные черты строения сходны с другими представителями подсемейства лентивирусов. Наружная оболочка вируса, или “конверт” состоит из бимолекулярного слоя липидов, который имеет происхождение из клеточной мембраны клетки хозяина. В эту мембрану встроены рецепторные образования, по виду напоминающие грибы. “Шляпка гриба” состоит из четырех молекул гликопротеида gp120, который обладает сродством к молекулам CD4. “Ножка гриба” состоит из четырех молекул гликопротеида gp41, которые встроены в мембрану. Так как мембрана имеет клеточное происхождение, то на ее поверхности и внутри нее сохраняется множество клеточных белков. Под наружной оболочкой располагается сердцевина вируса (кор), которая имеет форму усеченного конуса и образована белком р24. Промежуток между наружной вирусной мембраной и сердцевиной вируса заполнен матриксным белком р17. Внутри сердцевины располагаются две молекулы вирусной РНК, связанные с низкомолекулярными белками (р9 и р7) основного характера. Каждая молекула РНК содержит 9 генов ВИЧ. Три из них - gag, env и pol - являются структурными. Имеются также три регуляторных гена: tat, rev и nef, и три дополнительных гена: vpu, vpr и vif. Эти гены содержат информацию, необходимую для продукции белков, которые управляют способностью вируса инфицировать клетку, реплицироваться и вызывать заболевание. Концы каждой молекулы РНК содержат дублированную последовательность РНК, так называемый длинный концевой повтор - LTR. Участки LTR действуют как переключатели для управления процессом вирусной транскрипции, взаимодействуя с белками ВИЧ или с белками клетки хозяина. Кроме РНК там же находятся вирусные ферменты: обратная транскриптаза, состоящая из двух субъединиц - р64/53, протеаза - р22, эндонуклеаза (интеграза) - р31. Обратная транскриптаза осуществляет синтез вирусной ДНК с молекулы вирусной РНК. Эндонуклеаза производит встраивание вирусной ДНК в геном клетки хозяина, в результате чего образуется провирус. Протеаза участвует в “нарезании” предшественников вирусных белков при созревании новой вирусной частицы.
СТРОЕНИЕ ГЕНОМА И ЭКСПРЕССИЯ ГЕНОВ ВИЧ.
Важность исследования строения генома ВИЧ обусловлена тем, что в основе всех патологических процессов, происходящих при заражении вирусом, лежит экспрессия вирусных генов.
Изучение структуры генетического аппарата ВИЧ с помощью молекулярного клонирования выявило его сложную организацию и значительные различия между изолятами. ДНК провируса имеет 9283 пары нуклеотидов (п.н.) и окружена длинными концевыми повторами - LTR - в 638 п.н. В LTR выявляются все обычные регуляторные элементы. (см. Рисунок 1).
В качестве затравки при синтезе минус цепи ДНК ВИЧ используется тРНКлиз, в то время как большинство ретровирусов млекопитающих используют тРНКпро. ТРНКлиз используется и при синтезе минус-цепи ДНК вируса опухоли молочных желез мышей (MMTV), который имеет и очень сходную последовательность полипуринового тракта. Однако на этом сходство ВИЧ и MMTV заканчивается. Как стало ясно этот ретровирус имеет мало общего и с ретровирусами человека HTLV-1 и HTLV-2, хотя в ранних сообщениях говорилось об их взаимной гомологии. Наиболее близкими к ВИЧ как по морфологии, так и по течению вызываемого заболевания оказались вирусы группы лентивирусов. Клонированная провирусная ДНК вирусов висна и инфекционной анемии лошадей, относящихся к подсемейству лентивирусов, образует стабильные гибриды с провирусной ДНК ВИЧ. Анализ первичной последовательности нуклеотидов этих провирусных ДНК выявил обширные участки гомологии, особенно в областях генов gag и pol.
ГЕНЫ И БЕЛКИ ВИЧ.
gag. Первая открытая рамка кодирует внутренние белки вириона. Эти белки вместе с белками, кодируемыми геном pol, прочитываются, как и у других ретровирусов, с полноразмерной РНК в 9300 нуклеотидов. В результате трансляции этой иРНК (см. Рисунок 2) образуется предшественник с мол. Массой 55 кД. В процессе дальнейшего протеолитического расщепления этот белок нарезается на p17, p24, p9 и р7. Согласно наблюдениям, в сыворотках больных СПИД обнаруживаются антитела ко всем этим продуктам. Значительную фракцию составляют антитела к р24 - основному внутреннему белку вириона. Антитела к р24 обычно появляются на ранних стадиях заболевания и часто исчезают по мере его прогрессирования.
pol. Как и у других ретровирусов, кодируемые этим геном белки считываются в виде gag-pol предшественника. Поскольку рамка считывания гена pol не совпадает с рамкой gag, при созревании иРНКpol должно происходить удаление небольшого интрона, сдвигающее рамку считывания. Анализ первичной нуклеотидной последовательности области перекрывания генов gag и pol выявляет присутствие там нескольких участков, которые могут выполнять функцию акцепторных сайтов сплайсинга. Другим механизмом совмещения рамок считывания является так называемый “перескок рамки” при трансляции. В результате рибосомы “перепрыгивают” через стоп-кодон, ограничивающий рамку gag, и прочитывают pol уже в правильной рамке считывания. Подобный механизм описан для некоторых ретровирусов.
Ген pol кодирует 3 фермента: протеазу (р22), обратную транскриптазу (р64/53) и эндонуклеазу (р31). Эти белки образуются в результате протеолитического расщепления предшественника с молю массой 150кД. Несмотря на относительно небольшое количество этих белков в вирионе (примерно 2 молекулы на вирион), антитела к ним выявляются в сыворотках больных СПИДом. Наиболее ярко выражена реакция с р31.
sor .Tретья открытая рамка перекрывается с 3`-концом гена pol и кодирует белок с молю массой 23 кД. Антитела к этому белку удается выявить в сыворотках больных СПИДом. По-видимому, белок транслируется со сплайсированных полиаденилированных РНК размером 5500 и 5000 нуклеотидов (см. Рисунок 2). Как показали опыты с использованием инфекционной провирусной ДНК ВИЧ, мутации в области гена практически не влияли на способность вируса реплицироваться и оказывать цитопатогенное действие на CD4-клеточную линию, если не считать небольшого замедления этих процессов по сравнению с исходным вирусом. Тем не менее, высокая консервативность нуклеотидной последовательности гена sor указывает на наличие какой-то функции продукта этого гена в жизненном цикле вируса. Возможно, эта функция важна при репликации в нелимфоидных клетках, например в нервных и ретикулоэпителиальных.
env. иРНК, кодирующая белки оболочки вириона, образуется в результате сплайсинга, приводящего к удалению из геномной РНК большого интрона, содержащего гены gag, pol и sor (см. Рисунок 2). Образующаяся иРНК размером 4300 нуклеотида
содержит открытую рамку с типичным инициирующим AUG, которая может направлять синтез белка, состоящего из 861 аминокислотного остатка с мол. массой 97.5 кД. Этот белок предшественник в дальнейшем обильно гликозилируется, в результате чего его мол. масса возрастает до 160 кД. Предшественник содержит 3 гидрофобные области, характерные для оболочечных белков других ретровирусов. Первый гидрофобный участок (с 17-й по 31-ю аминокислоты) соответствует сигнальному пептиду, второй находится в районе сайта протеолитического расщепления белка-предшественника, третий является частью трансмембранного белка. В результате протеолитического расщепления образуется 2 сильно гликозилированных белка: наружный белок оболочки gp120 и трансмембранный белок gp41. Интересной особенностью трансмембранного белка является наличие необычно длинной последовательности (длиной в 150 аминокислотных остатков) гидрофильных аминокислот вслед за гидрофобной частью трансмембранного белка. Эта последовательность, по-видимому, является внутриклеточным фрагментом gp41. Еще, как показали исследования, правильный процессинг gp160 происходит не во всех клеточных линиях. От чего это зависит, пока неизвестно.
3`-orf. Эта открытая рамка расположена между 8347-м и 8992-м нуклеотидами и простирается, таким образом, в U3 область 3`-LTR. Кодируемый этим геном белок имеет мол. массу 27 кД и транслируется со сплайсированной иРНК размером 1800 нуклеотидов (см. Рисунок 2).
Хотя антитела к этому белку удается выявить в крови больных СПИДом, он не является абсолютно необходимым для репликации вируса. Продукт 3`-orf оказывает влияние на цитопатогенность вируса.
tat-3. Явление трансактивации было впервые описано для ретровирусов человека HTLV-1 и HTLV-2. Белок, осуществляющий функцию трансактивации, кодируется у этих вирусов небольшой открытой рамкой, расположенной на 3`-конце генома после гена env. Механизм его действия заключается в активации транскрипции структурных генов вируса, вследствие чего ген, кодирующий белок-трансактиватор, был назван tat (transactivator of transcription).
Феномен трансактивации выражен у ВИЧ на несколько порядков сильнее, чем у HTLV-1 и HTLV-2. Как сейчас стало ясно, за этот процесс у ВИЧ отвечают по крайней мере, 2 гена: tat-3 и art (trs). Первый из них кодируется иРНК размером около 2000 нуклеотидов, образующейся в результате сложного сплайсинга (см. Рисунок 2). Механизм действия белка tat-3 у ВИЧ значительно сложнее, чем у аналогичных белков tat вирусов HTLV-1 и HTLV-2.
Продукт гена tat-3 - белок с мол. массой 14 кД, выявляемый с помощью сывороток больных СПИД. Мутации в 5`-области первого кодирующего экзона tat-3 нарушают способность вируса синтезировать структурные белки и реплицироваться. Эти мутации могут быть комплементированы в клеточных линиях, постоянно экспрессирующих tat-3 белок. Сейчас получены линии как В- так и Т-лимфоцитов, стабильно трансформированных tat-3 геном и продуцирующих белок-трансактиватор. Другие клеточные линии, например HeLa, продуцирующие функциональный tat-3 белок, также могут поддерживать размножение мутантного по tat-3 ВИЧ. Использование подобных клеточных линий и клонированных провирусных ДНК, содержащих различного размера делеции в tat-3 гене, позволило изучить механизмы действия кодируемого этим геном белка.
art (trs). Другим белком, участвующим в регуляции экспрессии структурных генов ВИЧ является продукт гена art (antirepression transactivator - антирепрессорный трансактиватор). Транслируется он, по-видимому, с иРНК, принадлежащей к тому же классу молекул размером 2000 нуклеотидов, что и иРНКtat-3. Кодирующие экзоны гена art перекрываются экзонами tat-3 (см. Рисунок 2), а при сплайсинге используются те же акцепторные и донорные сайты. Однако при трансляции art функционирует инициаторный AUG с координатой 5500, а не 5412, как для гена tat-3. В результате, art читается со сдвигом рамки отностительно tat-3, что приводит к уменьшению ее кодирующей рамки в первом транслируемом экзоне с 214 до 76 нуклеотидов и к увеличению во втором с 44 до 271 нуклеотида. Синтезируемый белок состоит из 116 аминокислотных остатков, причем основная доля приходится на аминокислоты, проявляющие основные свойства. Подобные белки обладают сродством к нуклеиновым кислотам и часто регулируют экспрессию генов.
Действие продукта art осуществляется на посттранскрипционном уровне. По-видимому, он активирует трансляцию иРНК структурных генов gag и env, снимая действие специфических негативных регуляторов (см. ниже). Наряду с этим продукт art участвует и в регуляции сплайсинга РНК, в связи с чем для указанного гена было предложено другое название - trs (transregulator of splicing - трансрегулятор сплайсинга).
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ВИРУСНЫХ ГЕНОВ.
Как было отмечено выше, геном ВИЧ, помимо обычных ретровирусных генов (gag, pol, env), содержит еще ряд генов, функции которых заключаются в регуляции экспрессии синтеза структурных белков вириона. Если о функции продуктов генов sor и 3`-orf мало что известно, кроме того, что они не являются необходимыми для репликации вируса, о действии продуктов генов tat-3 и art имеется много данных, укладывающихся в определенную схему.
Первые опыты по изучению синтеза некоторых ферментов в клетках, трансформированных рекомбинантными ДНК, содержащими гены этих ферментов под контролем LTR ВИЧ, показали, что количество специфических иРНК резко увеличивается (в 100-1000 раз) после введения в клетки дополнительно активно экспрессирующегося tat-3 гена. Дальнейшие опыты показали, что продукт гена tat-3 увеличивает количество специфических иРНК примерно в 10 раз, причем его действие опосредовано последовательностью, присутствующей в R-элементе LTR ВИЧ. Однако для максимального эффекта tat-3-зависимой трансактивации необходимо присутствие последовательности U3 - элемента LTR. Таким образом, продукт гена tat-3 вызывает активацию транскрипции, и его активирующее действие осуществляется на этапе инициации транскрипции.
Важной частью tat-3-зависимой трансактивации является активация трансляции иРНК. Этот посттранскрипционный эффект tat-3-белка также опосредован последовательностью R-элемента LTR ВИЧ. Эта последовательность была обозначена TAR (trains acting responsive) и локализована между -17 и +80 нуклеотидами в LTR ВИЧ. Анализ первичной нуклеотидной последовательности этого участка показал наличие там 24-х нуклеотидного обращенного повтора (между 1-м и 59-м нуклеотидами), который может формировать петлю в иРНК. Такая вторичная структура иРНК может мешать 40S субъединице рибосомы передвигаться вдоль иРНК при инициации трансляции и препятствовать, таким образом, синтезу продукта. Взаимодействие продукта гена tat-3 с этим участком иРНК релаксирует структуру и снимает, таким образом, препятствие для трансляции. В эксперименте было выяснено, что делеции в TAR участке снимали тормозящее действие R-элемента на трансляцию, однако полностью ликвидировали эффект трансактивации. Таким образом, tat-3 кодируемый белок или индуцированные им клеточные факторы узнают TAR последовательность как в ДНК (провирус), так и в иРНК и активирует соответственно транскрипцию или трансляцию.
Действие другого гена-трансактиватора - art- осуществляется только на посттранскрипционном уровне. Продукт гена art(trs) участвует в регуляции сплайсинга. Он снимает репрессию специфических негативных регуляторов трансляции, расположенных на иРНК, кодирующей белки gag и env. В отсутствие активного гена art(trs) нарушается синтез именно этих белков, в то время как экспрессия других генов (например tat-3) не нарушается.
Если суммировать приведенные данные, то можно предложить такую модель регуляции экспрессии генов ВИЧ. Продукт гена tat-3, взаимодействуя с TAR последовательностью в LTR провируса, активирует транскрипцию провирусной ДНК. Возможно, это происходит за счет уменьшения тормозящего действия на процесс транскрипции специфического негативного регуляторного элемента (NRE), расположенного между -340-м и -185-м нуклеотидами в U3 последовательности LTR ВИЧ.
Тот же tat-3 белок активирует и трансляцию иРНК, снимая ингибирующее действие на инициацию трансляции TAR-элемента. Для полноценного синтеза структурных белков вириона необходимо действие еще одного белка - продукта гена art(trs). Этот белок появляется в результате описанных выше событий и переключает сплайсинг на производство иРНК структурных генов. Этот же белок снимает действие негативных регуляторов трансляции, расположенных на иРНК, кодирующих структурные белки (gag и env). Предполагаемая цепь событий представлена на рисунке 3.
Конечно, предполагаемая схема содержит много “белых пятен”. Так, пока не ясно, что является исходным толчком к активации провируса. Такая активация происходит при антигенной стимуляции зараженного ВИЧ Т-лимфоцита, однако молекулярные механизмы этого явления не изучены. Важную роль в активации провируса может играть и взаимодействие вирусных генов с клеточными белками. Так, фактор транскрипции Sp1, присутствующий в клетках млекопитающих, связывается с промоторным участком в LTR промотора ВИЧ и активирует синтез РНК в 5-10 раз.
ТЕОРИИ ПРОИСХОЖДЕНИЯ ВИЧ.
Вопрос о происхождении ВИЧ 1-го и 2-го типов является одним из ключевых в современном понимании проблемы СПИДа, поскольку его решение в значительной мере определяет направления диагностики, лечения и профилактики данной патологии.
Итак, известны следующие возбудители СПИД:
n вирус иммунодефицита человека(ВИЧ) - LAV/HTLV-3 (HIV, ВИЧ-1), распространенный в основном в Америке, Европе, центральных, восточных и южных районах Африки;
n родственные обезьяньим вирусы подгруппы западно-африканских Т-лимфотропных ретровирусов (LAV-2, HIV-2, ВИЧ-2), выделенные группой Л.Монтанье в 1986 г. От больных СПИД африканцев из Гвинеи.
n вирус HTLV-4, выделенный американскими учеными от представителей групп повышенного риска (Сенегал). Распространен в основном в Африке.
Описаны случаи одновременного существования в организме человека ВИЧ-1, ВИЧ-2 и HTLV-4 в различных сочетаниях.
По поводу происхождения вирусов иммунодефицита человека в литературе представлено много данных, порою противоречивых и не всегда полных. Споры продолжаются. По мере накопления материала возникли гипотезы происхождения ВИЧ, основные из которых будут приведены ниже.
Вирус создан искусственно в конце 70-х годов текущего столетия посредством методов генной инженерии на основе новых знаний о влиянии различного рода излучений, иммунодепрессантов и мононуклеарных антител на различные звенья иммунной системы.
В естественных условиях вирус иммунодефицита человека может иметь антропогенное происхождение, а именно:
* ВИЧ - типичный экзогенный ретровирус, существовавший у людей с древних времен и эволюционировавший вместе с человеком при его расселении на Земле;
* в глухих уголках Центральной Африки ВИЧ циркулировал и вызывал эндемические заболевания СПИД длительное время, затем через о. Гаити попал в США и в последующем довольно быстро распространялся на все континенты;
* ВИЧ - не африканского происхождения, а возник и до нынешней пандемии, существовал в странах умеренного климата (Северная Америка, Европа), в силу слабой патогенности вызывая отдельные заболевания, практически не диагностируемые как СПИД
Зоонозное происхождение вируса иммунодефицита человека, что могло быть реализовано следующими путями:
* в начале 50-х годов нашего столетия конструирование ВИЧ произошло при генетических рекомбинациях (видимо, случайных) вируса лейкоза человека и животных (ретровируса типа С) с вирусом опухоли молочной железы мышей (ретровирус типа В) или с вирусом обезьяньего СПИД (ретровирус типа D);
* в древние времена мутанты вируса иммунодефицита зеленой мартышки трансформировались и обрели нового хозяина - человека;
* по структуре генома и биологическим свойствам ВИЧ близок к лентивирусу висна и вирусу инфекционной анемии лошадей, отмечается выраженная общность их внутренних (сердцевинных) белков.
Ряд авторов утверждают, что пробел в структуре и свойствах между вирусами иммунодефицита обезьян и человека уже частично заполнен сходными вирусами HTLV-4, ВИЧ-2, а также недавно открытым шведским ученым вирусом SBL, и процесс этот будет продолжаться.
Однако проблема СПИДа сегодня представляется настолько нетривиальной и многоплановой, что традиционный эпидемиологический подход к указанному заболеванию как к обычной антропозоонозной инфекции вряд ли может быть исчерпывающим в трактовке генеза ВИЧ. В пояснение этому необходимо отметить, что в эволюционном аспекте ретровирусы (в первую очередь онкогенные) часто и абсолютно оправданно рассматриваются в неразрывной связи с мобильными клеточными элементами генома эукариот. Примечательным является факт удивительного структурного сходства провирусных нуклеотидных последовательностей и перемещающихся элементов эукариотических геномов.
В первую очередь обращает на себя внимание тот факт, что как проретровирусы, так и мобильные генетические элементы ограничены с обеих сторон регуляторными последовательностями - длинными концевыми повторами (LTR).
Более того, LTR проретровирусов и подвижных генетических элементов сами характеризуются сходством молекулярной организации. У всех перечисленных полинуклеотидных последовательностей LTR несут на концах инвертированные концевые повторы. Появление коротких концевых повторов по краям проретровирусов и перемещающихся элементов генома эукариот связано с удвоением соседних участков ДНК, то есть указывает на общий механизм их интеграции с клеточным геномом. Одной из общих характеристик процесса интеграции в геном мобильных элементов (ретропозонов) и ретровирусов является использование обратной транскрипции.
Сходным образом осуществляется и их прямая транскрипция: как провирусы, так и мобильные генетические элементы транскрибируются в виде полноразмерной РНК от начала одного LTR до конца другого. Оба вида образующихся транскриптов имеют почти одинаковую длину.
Приведенные выше и некоторые другие признаки структурно-функционального сходства ретровирусов и мобильных генетических элементов дали основания Х. Темину выдвинуть гипотезу о происхождении ретровирусов из мобильных клеточных элементов эукариотического генома. Согласно предположению Темина эволюция ретровирусов начиналась с того, что два небольших мобильных элемента, переместившись к краям гена ДНК-полимеразы, в дальнейшем в виде единого транспозона мигрировали совместно с названным геном, причем последний постепенно трансформировался в ген РНК-зависимой ДНК-полимеразы (обратной транскриптазы), а фланкирующие последовательности превратились в LTR. В дальнейшем подобный новообразованный мобильный элемент в процессе транспозиции мог захватить и включить в свой состав другие структурные гены, характерные для ретровирусов (gag, env).
Одним из убедительных современных подтверждений Темина могут служить данные, указывающие на высокую степень структурно-функциональной близости ретровирусов и мобильного элемента генома дрожжей Ту. Ретротранспозон Ту обладает длинными концевыми повторами и двумя структурными генами: Ту А и Ту В. При этом ген Ту А является аналогом гена gag ретровирусов, кодирующего синтез сердцевинных белков вириона, а Ту В - аналогом гена pol, управляющего синтезом обратной транскриптазы и протеазы.
Если признать правомочным вытекающее из изложенного предположения о происхождении ВИЧ из ретротранспозонов лимфоидных клеток человека, то следующим закономерным этапом было бы решение вопроса о механизмах активации определенных ретротранспозонов и приобретения ими автономности с последующим функционировании в форме ВИЧ.
Необходимо отметить, что идея связи иммунодефицитных состояний в целом с нестабильностью генома уже получила достаточно широкое распространение в клинических наблюдениях и экспериментальных исследованиях. Однако в качестве характеристики нестабильности генома в большей части проанализированных работ рассматривается только уровень хромосомных аберраций, являющийся показателем грубых нарушений структуры хроматина. Конкретные молекулярные механизмы лабилизации генома в упомянутых исследованиях не освещены.
Одним из таких принципиальных механизмов можно считать неспецифические реакции клеточного стресса, сопровождающиеся синтезом белков теплового шока. Убедительно показано, что тепловой шок и некоторые химические индукторы клеточного стресса, приводящие к экстренному синтезу стрессовых белков, резко активируют экспрессию длинных концевых повторов ВИЧ а культуре клеток. Более того, зона LTR ВИЧ, которая связывает белки, распознающие ДНК, оказывается гомологичной соответствующей зоне в составе генов синтеза белков теплового шока. Это указывает на отчетливую взаимосвязь экспрессии генетических элементов ретровируса (ретротранспозона?) с клеточным стрессом. Если подобные явления имеют место in vivo, то это позволило бы приблизиться к пониманию не только самих внутриклеточных процессов, но и природы конкретных биохимических медиаторов (типа белков теплового шока), способствующих увеличению лабильности генома.
Среди факторов-индукторов клеточного стресса особая роль принадлежит ультрафиолетовому излучению. Продемонстрирована интенсивная наработка мРНК, гибридизующейся с ДНК гена белка теплового шока hsp 70, под влиянием ультрафиолета in vivo.
Следовательно, УФ, индуцируя синтез белков теплового шока в облученных клетках, должно было бы тем самым усиливать экспрессию элементов генома ВИЧ. Действительно, в экспериментах установлен факт более чем 150-кратного увеличения экспрессии ВИЧ-специфичных LTR, интегрированных в геном клеток HeLa, после облучения их УФ. Более того, степень ее интенсификации оказалась пропорциональной дозе излучения.
При анализе результатов исследований постоянно возникал вопрос о конкретных механизмах действия УФ-излучения как фактора геномного стресса, приводящего к активации экспрессии LTR ВИЧ. Особое внимание привлекла гипотеза участия в названном процессе УФ-эндонуклеазы - фермента, катализирующего образование однонитевых разрывов ДНК под влиянием УФ-излучения и инициирующего ее репарацию.
После ряда экспериментов для подтверждения данной гипотезы был сделан вывод, что лимфоциты крови больных ВИЧ-инфекцией характеризуются повышенной лабильностью генома, степень которого возрастает по мере развития заболевания. Одним из ключевых патобиохимических механизмов дестабилизации генома лимфоцитов в этих условиях являются процессы свободнорадикального окисления. Атака ДНК лимфоцитов свободными радикалами вызывает образование однонитевых разрывов, что одновременно служит инициирующим сигналом для репарации указанных повреждений.
УФ-эндонуклеаза, активируемая под влиянием УФ-миметического действия активных форм кислорода, инициирует репарационные процессы, которые в силу своей избыточной интенсивности приводят к формированию олигонуклеотидных последовательностей гораздо большей протяженности, чем это необходимо для репарации разрывов ДНК. Возможно, что образующиеся последовательности могут служить либо факторами регуляции продукции и экспрессии ретротранспозонов (в зависимости от сайтов их встраивания в репарирующую ДНК), либо даже структурными их элементами. В пользу данного предположения косвенным образом свидетельствуют результаты работы, согласно которым тепловой шок и окислительный стресс существенно усиливают транскрипцию ретротранспозонов у дрозофилы.
Последующая автономизация ретротранспозонов в лимфоидных клетках (в том числе в процессе пассирования через организм реципиента генетического материала лимфоцитов) посредством соответствующих реаранжировок и сплайсинга, согласно представленной гипотезе, приводит к постепенной трансформации ретротранспозонов в ВИЧ.
Несомненно, значение факта значительной лабилизации генома лимфоцитов в условиях ВИЧ-инфекции может быть интерпретировано неоднозначно. Можно предположить, что дестабилизированный геном лимфоидной клетки является гораздо более восприимчивым к интеграции в него ДНК-транскриптов ВИЧ и, таким образом, подобные лимфоциты являются лишь “благодатной почвой” для распространения ВИЧ-инфекции. Однако имеется ряд результатов, которые могут свидетельствовать в пользу предложенной выше гипотезы.
При обследовании семи пациентов, в клетках которых отсутствовал генетический материал ВИЧ (по данным исследования методом ДНК-зондов), а в сыворотке отсутствовали антитела к белкам ВИЧ, было установлено, что после переливания донорской крови (не имевшей никаких маркеров ВИЧ) и последующей процедуры реинфузии аутокрови, облученной ультрафиолетом, в сроки от 1 до 3 месяцев в сыворотке этих пациентов появляются антитела к белкам р17 и р24 ВИЧ. Результаты обнаружения провирусных последовательностей с помощью ДНК-зондов в лимфоцитах обследованных к настоящему моменту оцениваются как неопределенные.
Безусловно, эти данные еще трудно трактовать как однозначные, однако в целом они могут являться подтверждением положения: проблема СПИДа как в фундаментальных, так и в прикладных ее аспектах не может быть решена в рамках традиционных подходов к рассматриваемой патологии как к тривиальной вирусной инфекции.
Очевидно, что многие из имеющихся сведений и гипотез о природе возбудителя синдрома приобретенного иммунодефицита человека требуют уточнения, дополнительных исследований и всестороннего анализа.
ПЕРЕДАЧА ВИЧ-ИНФЕКЦИИ.
ВИЧ передается типичным для всех ретровирусов способами, то есть “вертикально” (ребенку от матери) и “горизонтально”, особенно при половых контактах. Кроме этого к “горизонтальному” пути передачи относятся парентеральный - инъекционный, при попадании любой биологической жидкости от ВИЧ-инфицированного на кожу и слизистые при наличии на них микротравм. Следует, правда, отметить, что в разных биологических жидкостях титр вируса может быть очень различен: максимальная концентрация наблюдается в сперме и крови, а, например, в слюне или слезной жидкости могут находиться лишь единичные экземпляры вируса. Еще одно обстоятельство влияющее на передачу инфекции: некоторые носители ВИЧ выделяют намного больше вируса чем другие, хотя титр вируса в зараженном материале измерить довольно трудно. Передача вируса определяется также другими факторами, такими как травмы, вторичные инфекции, эффективность эпителиальных барьеров, а также присутствие или отсутствие клеток с рецепторами для ВИЧ.. Эти факторы позволяют объяснить, почему при одних обстоятельствах, например, при гомосексуальных контактах, вирус распространяется быстрее, чем при других.
Важным фактором, влияющим на заразность, может быть стадия инфекции. При большинстве вирусных инфекций наивысшие титры вируса достигаются на ранних стадиях, еще до образования антител. В случае ВИЧ эту фазу трудно изучать, поскольку она обычно бессимптомна, а гуморальный противовирусный ответ еще слаб или вообще не выявляется. Тем не менее, эта стадия, по-видимому, наиболее опасна для окружающих. Есть также данные о том, что больной вновь становится более инфекционным, когда уже развивается СПИД.
Суммируя все вышесказанное, можно выделить следующие факторы, влияющие на передачу ВИЧ-инфекции:
* титр вируса
* травма
* вторичная инфекция
* эпителиальные рецепторы
* интенсивность экспозиции
* фаза инфекции
КО-ФАКТОРЫ ВИЧ-ИНФЕКЦИИ.
В пандемии ВИЧ-инфекции велика роль ко-факторов, то есть факторов, способствующих ее распространению. Они либо активизируют те или иные пути инфицирования, либо повышают чувствительность организма к заражению.
Один из важных ко-факторов - число половых партнеров. Гомосексуалисты, больные СПИДом, имеют в среднем в течение жизни около 1100 половых партнеров, здоровые гомосексуалисты - 500, гетеросексуалисты -25.
Облегчают распространение эпидемии и венерические заболевания. Через появляющиеся при них изъязвления и другие нарушения целостности слизистых оболочек и кожных покровов половых органов проникает вирус СПИДа.
Различные вирусные инфекции ослабляют, а то и сводят на нет иммунитет организма. В первую очередь речь идет о вирусном гепатите В. В мире насчитывается примерно 200 миллионов носителей этого вируса. Им ежегодно заражаются 10-15 процентов гомосексуалистов - многие из них хронически инфицированы. Резко повышают чувствительность к ВИЧ такие инфекции, как туберкулез. Гомосексуалисты обычно предрасположены к иммунодепресси из-за систематического поступления в организм спермы, которая обладает выраженным иммунодепрессивным действием.
К ко-факторам относиться также возраст: 90% больных СПИДом заболевает в возрасте между 20 и 45 годами. СПИД - причина смерти мужчин в промежутке между 25 и 44 годами, что связано с большой сексуальной активностью и наркоманией именно в этот период.
Более злокачественное течение СПИДа у детей объясняется незрелостью их иммунной системы.
Повторные инфекции и переливания крови у больных гемофилией стимулируют иммунную систему, что тоже благоприятствует обострению чувствительности этих лиц к ВИЧ.
Ко-факторами могут быть и вирусы, которые недавно обнаружили у больных СПИДом. Ученые из Института патологии Вооруженных сил США выделили из организма 23 из 24 больных СПИДом ранее неизвестный вирус. Предполагается, что кооперация этого вируса с ВИЧ ведет к саркоме Капоши, которая часто развивается при СПИДе.
Р.Галло с сотрудниками открыли новый вирус, относящийся к группе герпеса. Его выявили в клетках больных с лимфоаденопатией, которые были инфицированы ВИЧ. Вирус был назван NBLV (человеческий В-лимфотропный вирус). Вполне возможно, что и он - ко-фактор ВИЧ и “участвует” в образовании опухолей лимфоидной ткани.
ПАТОГЕНЕЗ И КЛИНИКА ВИЧ-ИНФЕКЦИИ.
ПАТОГЕНЕЗ. У разных индивидуумов “отзывчивость” организма на заражение ВИЧ неодинакова. Это зависит от специфики их системы гистосовместимости. У ВИЧ-инфицированных чаще, чем у других, встречается HLA-B35 фенотип. Установлено, что тот же фенотип - главный фактор риска для заражения ВИЧ наркоманов, вводящих наркотики внутривенно.
В организме вирус внедряется в чувствительные клетки. Основные клетки мишени CD4-лимфоциты (хелперы), на их поверхности есть молекулы CD4-рецепторы, способные связываться с поверхностным белком ВИЧ - gp120. В меньшем числе они содержаться на мембранах макрофагов, еще в меньшем на мембранах В-лимфоцитов. Кроме того, ВИЧ проникает в ЦНС, поражая нервные клетки и клетки нейроглии, в хромаффинные клетки кишечника, в сперму.
МЕХАНИЗМЫ ВЗАИМОДЕЙСТВИЯ ВИЧ С РАЗЛИЧНЫМИ ЗВЕНЬЯМИ ИММУННОЙ СИСТЕМЫ
Известно, что продуктивное взаимодействие вируса и клетки включает несколько фаз: адсорбцию вируса и его проникновение в клетку, “раздевание” вируса, биосинтез вирусных компонентов, формирование вирусных частиц и выход вируса из клетки. Вирус существенно изменяет метаболизм клетки, и трансформированная клетка уже по особому взаимодействует как с родственными, так и с другими клетками организма
Начало заболевания обычно проявляется на фоне интенсивной репродукции вирусов в клетках РЭС, эпителии дыхательной или пищеварительной систем, крови и др. Возникающие в результате иммунного ответа антитела в определенной степени препятствуют распространению вируса в организме, но существенно не влияют на процессы его репродукции. Создается впечатление, что антитела не только не определяют исход первичной вирусной инфекции, а более того, могут маскировать циркулирующий вирус в крови и других жидкостях организма. Более выражены в начальном периоде инфицирования факторы неспецифической защиты - повышенная температура, повышение уровня различных ингибиторов, интерферона и др.
В патогенезе СПИД, по мнению большинства исследователей, решающее значение имеет избирательное цитопатическое воздействие вируса СПИД на Т-лимфоциты-хелперы-индукторы, что в первую очередь проявляется в снижении противовирусного, противомикробного и противоопухолевого иммунитета. Следствием указанного является накопление вируса в крови и интенсивное его распространение в организме, усугубление течения и необратимое развитие оппортунистических инфекций, ускорение злокачественного роста вследствие преморбидного действия персистирующего в клетках опухоли частиц вируса и поражения натуральных киллеров. Первичное селективное цитодеструктивное действие вируса на Т-хелперы-индукторы проявляется в развитии лимфопении, снижении числа циркулирующих в крови Т4-клеток, нарушении соотношения Т4/Т8 и угнетение пролиферации Т-лимфоцитов в ответ на действие митогенов (фитогемагглютинин, конкавалин А, специфические антигены), снижении или полном отсутствии реакции кожной гиперчувствительности к кожным аллергенам, снижении пролиферации аутологичных лимфоцитов в смешанной реакции, уменьшение продукции гамма-глобулина лейкоцитами больных оппортунистическими инфекциями, ослабление экспресси Т-клеточных рецепторов к интерлейкину-2, дефектности продукции лимфокинов, поликлональной активации В-лимфоцитов с продукцией большого числа фракций гамма-глобулина.
Иммунологические нарушения при СПИД проявляются также в повышении уровня циркулирующих иммунных комплексов, антилимфоцитарных антител невыясненной специфичности, в увеличении количества и лабильности альфа-интерферона. Однако описанные выше иммунологические аномалии не являются строго специфичными именно для СПИД , они встречаются и при иммунодефицитах другой этиологии; дифференциация специфичности поражения особенно затруднительна в начальный период заболевания.
Возбудитель СПИД имеет выраженный тропизм к Т-лимфоцитам и другим клеткам с антигенной формулой - ОКТ-4 (Т-хелперы, В-клетки, моноциты, макрофаги). Указанное сродство обусловлено чрезвычайно высоким аффинитетом гликопротеида наружной оболочки вириона к рецептору Т4 (CD4), располагающемуся на поверхности Т-хелперов. Было доказано, что мембранный белок лимфоцитов CD4 служит рецептором для вируса СПИД. Что касается вирусных белков, то лишь один из них с молекулярной массой 11000 Д (белок gp120) способен связываться с CD4 белком.
Для ряда вирусов человека установлена связь клеточной чувствительности к ним с определенными хромосомами человеческих клеток. Например, в хромосоме19 локализованы гены чувствительности к вирусам полиомиелита, в хромосоме 3 - к вирусу герпеса, в хромосоме 21 - к вирусам Коксаки В. В какой хромосоме существуют гены чувствительности к вирусу СПИД, пока не установлено.
Генетические исследования показали связь развития СПИД с определенным типом антигенов главного комплекса гистосовместимости (МНС). Установлено, что среди больных СПИД весьма значительно количество лиц, имеющих HLA-DR5 антигены. Подтверждено, что лица с таким фенотипом лимфоцитов составляют группу с повышенным риском заболевания саркомой Капоши, встречающейся в 6 раз чаще, чем в других группах.
Известно, что иммунореактивность зависит как от структуры антигена, так и от генетических особенностей организма и связана с одним из типов клеток или их субпопуляций, участвующих в иммунных реакциях. У слабо реагирующего организма нет Ir-гена (или он по какой-либо причине не реализует свои функции), позволяющего Т-хелперам распознать антиген и оказать помощь В-лимфоцитам. Недостаточность иммунного ответа на уровне В-клеток нередко обусловлена отсутствием другого гена, контролирующего синтез веществ клеточной поверхности (акцепторов), которые воспринимают дополнительный сигнал от Т-хелперов (Leu 3a+). Цепь событий в иммунной системе развертывается следующим образом. Антиген, захваченный макрофагом, разрушается до отдельных фрагментов гидролитическими ферментами. Затем в работу вступают Ir-гены, которые контролируют образование семейства так называемых Ia-белков. У разных индивдуумов или линий животных эти белки отличаются друг от друга по структуре, отражая различие в генах, которые обеспечивают их синтез. Если белки макрофагов способны вступать во взаимодействие с данным антигеном, то образовавшийся комплекс антиген-белок мигрирует к мембране макрофага и оказывается на ее поверхности. В результате создаются условия для работы Т-хелперов. Т-хелперы могут наиболее четко распознавать антиген только в ассоциации его с Ia-белком. Идентифицировав антиген, измененный Ia-белком, Т-хелперы оказывают необходимую помощь В-клеткам, которые без этого не приступают к синтезу иммуноглобулинов. Помощниками Т-В-кооперации служат также различные белки регуляторы, лимфокины, секретируемые Т-клетками.
Обнаружены антитела, нейтрализующие вирус СПИД, у больных со СПИД и СПИД-комплексом..
В последнее время установлено, что ВИЧ в иммунной системе инфицирует прежде всего макрофаги и моноциты, с помощью которых и распространяется в организме. Местом сохранения вполне могут быть также тромбоциты, В-лимфоциты, эпителиальные клетки, глиальные клетки нервной системы.
Антиген встречается с клетками Лангерганса, которые представляют его (в комплексе с антигеном гистосовместимости) Т-клеткам, запрограммированным отвечать на данный антиген ( подробнее о клетках Лангерганса ниже). Активированные таким образом клетки экспрессируют рецепторы к интерлейкину-1 (ИЛ-1). ИЛ-1 кроме лимфоидных клеток имеет и ряд иных клеток-мишеней, что обуславливает множественную направленность биологической активности этого вещества и его участие в целом ряде процессов, не относящихся собственно к иммунным. Выделяемый кератиноцитами (и, возможно, клетками Лангерганса) фактор заставляет активированные Т-клетки секретировать интерлейкин-2 (ИЛ-2) - важнейший тип лимфокина. Рецептором для ИЛ-2 на клетке является структура носящая название Tac-антигена. ИЛ-2 вступает в связь с рецепторами на поверхности других Т-клеток, отвечающих на данный антиген и находящихся в дерме, лимфатических узлах, селезенке, и инициирует их пролиферацию. Результатом указанного является новая популяция Т-клеток, вырабатывающих антитела к данному антигену.
ВЛИЯНИЕ ВИЧ НА Т-ХЕЛПЕРЫ
Существует целый ряд дополняющих друг друга механизмов взаимодействия ВИЧ с клетками Т-хелперов. Последние стимулируют работу Т-киллеров и макрофагов, индуцируют продукцию антител В-лимфоцитами. Лимфоциты-носители молекул Т8 (CD8) могут быть либо киллерами, либо супрессорами. Некоторые из механизмов еще спорны, подвергаются изменениям; постоянно появляются новые детали и нюансы влияния ВИЧ на иммунную систему
1-й механизм. При продуктивном процессе состоявшейся репродукции и массированном (несколько тысяч вирионов в генерации одной клетки) выходе из лимфоцитов ВИЧ интенсивно лизирует Т-хелперы. Но даже если вирусы спонтанно отпочковываются от Т-клетки-хелпера (без ее лизиса), клетка не успевает восстанавливать целостность мембран, молекулы цитоплазмы свободно элиминируют из клетки, и Т-хелпер гибнет. Поскольку Т-хелперы составляют порядка 60% циркулирующих Т-клеток, быстрая их гибель приводит к глубоким нарушениям иммунной системы инфицированного человека. СПИД развивается на фоне острой недостаточности CD4-лимфоцитов.
2-й механизм. На фоне общей стимуляции метаболизма лимфоцитов после их инфицирования вирусом, приводящей их к “гибели от истощения”, происходит интеграция геномов вируса и клетки. Диссиминация инфекции захватывает значительное число хелперных Т-лимфоцитов CD4+ фенотипа, при этом хронические инфекции и ряд других воздействий ведут к дополнительной стимуляции CD4+ популяции. Усиленная пролиферация Т-клеток ведет к активации супрессорных механизмов, увеличению количества CD8-лимфоцитов (Leu2a+ - Т-супрессоры) и резкому повышению их функции.
3-й механизм. Частицы ВИЧ изменяют реактогенные зоны поверхности Т-хелперов, что приводит к образованию нежизнеспособных синцитиев. Компоненты вирусной оболочки, синтезируемые в процессе репродукции вируса, резко нарушают цитоплазматическую мембрану клетки-хозяина: в результате элиминации протоплазмы клетки сливаются, образуются нежизнеспособные многоядерные структуры
Исследования подтвердили, что вирус резко изменяет мембраны Т-лимфоцитов и приводит к их слиянию в нежизнеспособные многоядерные клетки-монстры. Образование синцитиев возможно по типу гемагглютинирующего эффекта, когда здоровые лимфоциты при соприкосновении с инфицированными в свою оболочку включают поверхностные белки вируса, и взаимодействие рецепторных зон приводит к образованию крупных нежизнеспособных конгломератов.
4-й механизм. ВИЧ не разрушает CD4-лимфоциты, а изменяет и значительно замедляет их рост в периодической культуре, тогда как другие виды Т-клеток продолжают размножаться нормально. Отмечено, что скорость гибели зараженных клеток пропорциональна количеству CD4-рецепторов на их поверхности. Со временем число CD4-клеток становиться меньше, хотя некоторая их часть выживает и сохраняет вирус в латентном состоянии в виде провируса.
5-й механизм. ВИЧ маскирует CD4-маркер. Было показано, что в выживших CD4-лимфоцитах вирус может маскировать CD4-маркер на поверхности клеток или предотвращать его появление там. В результате получается, что число CD4-клеток еще меньше, чем на самом деле. С исчезновением CD4-клеток падает уровень ИЛ-2 и в результате замедляется рост клонов зрелых Т-клеток, индуцируемых этим лимфокином. Из-за ослабления синтеза интерлейкина и интерферона падает активность К-клеток и макрофагов, которые в номе стимулируютя этими белками.
В инфицированных ВИЧ клетках происходит снижение белков МНС класса 1 на их поверхности. Поскольку цитотоксические CD8-лимфоциты могут связывать антиген только вместе с белками МНС класса 1, этот эффект препятствует узнаванию и разрушению инфицированных вирусом клеток. Таким путем ВИЧ избегает любых воздействий со стороны иммунной системы, то есть создается ситуация “иммунного паралича”.
Антигены 1 класса главного комплекса гистосовместимости - молекулы HLA-A,B,C и 2 класса - HLA-DR- молекулы, имеющиеся на поверхности макрофагов, В-лимфоцитов, активированных Т-лимфоцитов. Молекулы 1 класса необходимы для распознавания поверхностных клеточных антигенов, а 2 класса - для контроля ответа. ВИЧ превращает Т-хелпер в донора супрессивного фактора. Р.Галло было сообщено, что ВИЧ вызывает не только уменьшение числа CD4-клеток и выделение растворимого фактора супрессии оставшимися лимфоцитами, но и делает эти уцелевшие клетки не способными осуществлять первую решающую стадию иммунного ответа - узнавание антигена. Это можно объяснить тем, что вирус вызывает повреждение рецепторов антигена на поверхности CD4-клеток. Этот рецептор подобен замку: для того, чтобы начался Т-клеточный ответ в него должен быть вставлен “ключ”, которым служит сочетание антигена и белка МНС класса. Возможно также, что вирус кодирует белок, попадающий на поверхность инфицированной клетки и препятствующий нормальной рецепции.
6-й механизм. В инфицированных CD4-клетках ВИЧ вызывает секрецию растворимого фактора супрессии. Это вещество блокирует иммунные реакции, зависящие от Т-клеток как in vitro, так и in vivo. При этом угнетается образование специфических антител и пролиферация Т-клеток.
Считают, что геном вируса не кодирует последовательность растворимых факторов супрессии, а только индуцирует в CD4-клеткет его синтез. Такой же механизм, возможно, лежит в основе иммуносупрессии при других инфекциях. Интересно было бы сравнить этот супрессивный фактор с иммуносупрессивным фактором продуктов перекисного окисления липидов.
7-й механизм. Вирус иммунодефицита человека вызывает изменения поверхности CD4-лимфоцитов, что провоцирует их уничтожение как чуждых иммунной системе. CD4-клетки, будучи инфицированными, погибают от того, что на них нападают Т-лимфоциты-киллеры. Таким путем идет непрерывное снижение количества Т-хелперов в крови, лимфоузлах, селезенке и других тканях. В то же время количество супрессорных CD8-лимфоцитов не уменьшается и даже несколько возрастает, что приводит к снижению показателя Тх/Тс.
8-й механизм. Проникший в лимфоциты ВИЧ изменяет геном Т-хелперов, в результате чего они лишаются способности к трансформации и нормальному ответу на ИЛ-2.
ВЗАИМОДЕЙСТВИЕ ВИЧ С Т-СУПРЕССОРАМИ
Дж.Леви с сотрудниками Калифорнийского университета установили, что иммунная система организма способна контролировать размножение вируса СПИД в зараженных клетках, а Т-супрессоры подавлять размножение ВИЧ. Учитывая, что только цитотоксические клетки и Т-супрессоры способна подавлять размножение ВИЧ, а их рецепторы устроены таким образом, что антиген связывается лишь одновременно с белками МНС класса1, в отличие от Т-хелперов, связывающихся с белками МНС класса 2, можно сделать заключение о возможном повреждении ВИЧ белков МНС класса 2.
РЕЗУЛЬТАТЫ ВЗАИМОДЕЙСТВИЯ ВИЧ С В-КЛЕТКАМИ
Кроме Т-клеток при ВИЧ-инфекции нарушается иммунорегуляторная роль и других субпопуляций лимфоцитов. Несмотря на нормальное количество циркулирующих В-лимфоцитов, реакция их на Т-зависимые антигены резко снижена, как понижены и ответы на Т-зависимые и независимые поликлональные активаторы. В то же время увеличено число В-лимфоцитов спонтанно секретирующих иммуноглобулины.
Сотрудники Института Аллергии и Инфекционных Заболеваний в Бетесде (Bethesda) установили, что ВИЧ непосредственно без посредничества Т-клеток, активирует В-лимфоциты. Пик интенсивности созревания и дифференцировки В-лимфоцитов отмечается на четвертые сутки с момента инкубации их с вирусом HIV, а в обычных условиях на активацию В-лимфоцитов Т-клетками уходит значительно больше времени. Максимум выделения антител приходится на 10-е сутки.
У больных СПИД повышен уровень IgA и IgG. Уровень IgM не изменен, в то же время повышен уровень острофазных белков, например, С-реактивного белка. Поскольку у В-лимфоцитов отсутствуют рецепторы CD4, а вирус все же сорбируется на их поверхности, вероятно, что ретровирус HIV способен взаимодействовать и с другими рецепторами или рецепторными полями иммунокомпетентных клеток.
МАКРОФАГИ И ВИЧ
В ряде работ убедительно показано, что эпидермальные макрофаги (клетки Лангерганса) “представляют” антиген таким образом, что преимущественно активируется цикл Т-хелперов. Если эти клетки вывести из иммунной игры (например, убив большой дозой УФ-облучения, радиацией), или антиген каким-либо путем минует их, в организме может возникнуть специфический иммунный “паралич”. Подобное развитие событий возможно и при ВИЧ-инфекции. Другие “представляющие клетки”, например дендритные клетки Грэнстейна, непосредственно стимулируют активность специфических Т-супрессоров, что дополняет высказанной предположение.
Известно, что лишь наличие трех клеточных типов (Т и В-клеток и макрофагов) в кооперации индуцирует полноценный иммунный ответ. Какая-либо одна или две из указанных популяций клеток не способны дать стимул к антителообразованию. При ВИЧ-инфекции скорее всего неполноценны два звена - Т-хелперное и макрофагальное.
При ВИЧ-инфекции из клеток РЭС макрофаги поражаются в первую очередь. Будучи инфицированы вирусом, эти клетки погибают не так быстро, как лимфоциты. Это, возможно, объясняется малым количеством рецепторов CD4 на их мембране.
Исходя из предположения, что ВИЧ тропен к одной из популяций макрофагов, а именно - к клеткам Лангерганса, проследим за сдвигами в иммунной системе инфицированного ВИЧ. Наличие в крови таких людей достаточно высокого уровня антител к вирусу иммунодефицита человека свидетельствует о том, что Ir-гены макрофага реализуют в полной мере свою функцию, в то время как низкие концентрации специфических антител объясняются поражением макрофагов. Поражение макрофагов отдельных органов (костного мозга, селезенки, лимфоузлов, легких), а также гистиоцитов соединительной ткани, микроглии, остеокластов, перитонеальных и плевральных макрофагов, звездчатых ретикулоцитов печени обуславливает направленность преимущественного поражения того или иного органа или ткани, что проявляется в клиническом течении СПИДа и фазах его развития. В то же время взаимодействие ВИЧ с клетками Лангерганса и Грэнстейна предшествует локальным поражениям других популяций макрофагов и является фоном для более агрессивного проявления Т-киллеров, предопределяющих гибель клеток-мишеней.
Взаимодействие ВИЧ с клетками Лангерганса и Грэнстейна интересно и с другой стороны. Когда Ia-белок в силу структурных особенностей антигена не может образовать с ним комплекс, последний оказывается на поверхности макрофага в свободной форме. Такой антиген кооперируется с Т-супрессором; в результате наблюдается подавление или полное отсутствие иммунного ответа. Известно, что супрессорные клетки могут связывать антиген с помощью молекул, сходных с иммуноглобулинами.
Подобно другим макрофагам, клетки Лангерганса несут рецепторы для Fc и C3. Благодаря их наличию клетки Лангерганса могут проявлять себя в качестве клеток-киллеров. Если это так, то вполне объяснимы весьма частые опухолевые процессы при СПИДе, так как клетки новообразований в первую очередь обследуются “киллерами”.
ВИЧ, попав в организм, начинает “представляться” макрофагам различных популяций, Т-киллерам, Т-хелперам, Т-супрессорам, В-клеткам организма. Изменение числа и функций клеток Лангерганса приводит к нарушениям в иммунологической цепочке “макрофаг (клетка Лангерганса) - Т-хелпер - В-клетка”. Во многих работах прямо или косвенно утверждается, что Т-хелпер, в основном, получает антиген от клетки Лангерганса, а антиген Т-супрессорам, в основном, представляет клетка Грэнстейна. Поэтому незрелые Т-хелперы не могут активизироваться, созреть, размножиться и (так как их число сократится по сравнению с количеством активированных, созревших и размножившихся при этом Т-киллеров и Т-супрессоров, получивших от “своих” макрофагов антиген) активизировать В-клетку. А если учесть уменьшение популяции Т-хелперов, инфицированных ВИЧ в числе первых при парентеральном заражении или получивших ВИЧ от зараженных клеток Лангерганса (при прочих, включая парентеральный способах заражения), то становиться очевидным, что число Т-клеток-хелперов вскоре станет заметно меньшим , так как зараженные ВИЧ клетки уничтожаются “киллерами”. Отсюда и все нарушения в поведении В-клетки. Поскольку она может и должна отреагировать на вирус СПИДа в крови, наряду с небольшим количеством антител против ВИЧ появляется увеличенное число спонтанно секретируемых В-лимфоцитами иммуноглобулинов. Так как снижен “союз” с Т-хелперами, несмотря на нормальное количество циркулирующих В-лимфоцитов, реакция их на Т-зависимые и Т-независимые поликлональные активаторы резко снижена. При этом само по себе проникновение “непредставленного” макрофагом антигена в лимфоузлы не индуцирует иммунитет, при этом не возникают клетки памяти.
Представленная картина нарушений при ВИЧ-инфекции, вызванная тропностью вируса к клеткам Лангерганса, очень близка к механизмам иммунных нарушений “болезни Т-хелперов”. Не исключая последнего, то есть что при СПИДе гибнут прежде всего Т-хелперы, возможно, что все же большая роль принадлежит клеткам Лангерганса как первичным мишеням и резервуарам накопления ВИЧ и последующего заражения Т-хелперов ( с одной клеткой Лангерганса могут одновременно контактировать от 1 до 7 Т-хелперов). По видимому, под воздействием ВИЧ в организме появляются или вещества угнетающие данные клетки ( по типу кортикостероидов) или же при дальнейшей активации очередной инфекцией клетки Лангерганса подвергаются ускоренной гибели, так как , отреагировав на любой новый антиген, клетка Лангерганса обязательно “представит” на своей поверхности большую дозу CD4 рецепторов, необходимых ей для связи с Т-хелперами.
ВЗАИМОДЕЙСТВИЕ ВИЧ С МОНОЦИТАМИ
Моноциты под действием ВИЧ гибнут медленнее, чем CD4-лимфоциты.В зараженных клетках вирус способен размножаться (резервуар вируса в организме), что приводит либо к их гибели, либо к уменьшению количества выделяемых ими факторов роста CD4-лимфоцитов. Моноциты больных СПИДом оказались способными продуцировать растворимый супрессивный фактор, подавляющий ответ Т- и В-лимфоцитов на митогены. Этот фактор зависит от Т-клеток и , видимо, связан с увеличением супрессирующего влияния со стороны последних. При СПИДе антимикробные функции моноцитов сохраняются, но они теряют способность к хемотаксису, падает их цитотоксическая активность и способность к продукции интерлейкина-1.
ВЗАИМОДЕЙСТВИЕ ВИЧ С ДРУГИМИ КЛЕТКАМИ ОРГАНИЗМА .
Помимо клеток крови резервуаром вируса могут быть клетки эндотелия кровеносных и лимфатических сосудов, эпителиальные клетки кожи и родственных тканей, глиальные клетки нервной системы и нервные клетки. Обычные при СПИДе психозы и атрофия мозга развиваются именно из-за способности вируса инфицировать ЦНС. Клетки за пределами сосудистого русла не всегда содержат поверхностные рецепторные белки к вирусу, что не позволяет ему адсорбироваться и проникать в клетку непосредственно, однако инфицирование может произойти при слиянии с зараженными CD4-клетками и макрофагами.
Проникая через гематоэнцефалический барьер, вирус вызывает в головном мозгу аномальное размножение глиальных клеток, окружающих нейроны, а также поражения, наступающие в результате потери белого вещества мозга, развития первичной лимфомы мозга и атрофии некоторых нервов (зрительного в первую очередь).
Поражение нервной системы отмечается у трети больных СПИДом и проявляется в четырех основных клинических формах:
* абсцессы мозга (прогрессирующая многоочаговая лейкоэнцефалопатия, криптококковый менингит, подострый энцефалит, возможно, цитомегаловирусной этиологии.);
* опухоли мозга, например первичная и вторичная лимфомы головного мозга;
* поражения сосудов мозга (церебральные геморрагии) и сердца (небактериальный тромботический эндокардит);
* очаговые поражения оболочек мозга, характеризующиеся самоограничиващимся менингитом.
СИСТЕМА ИНТЕРФЕРОНА ПРИ ВИЧ-ИНФЕКЦИИ.
Система интерферона (ИФ) является интегральной частью иммунной системы и обеспечивает координацию пролиферации, дифференцировки и активации эффекторных клеток иммунитета. В процессе иммунного ответа ИФ выполняет роль короткодистантных медиаторов межклеточных взаимодействий. ИФ определяет эффективность иммунного распознавания антигенов, влияя на экспрессию антигенов главного комплекса гистосовместимости 1-го и 2-го классов, а также карциноэмбриональных и опухолевых антигенов. ИФ играют определяющую роль в процессах элиминации антигенно-измененных “ своих” и чужеродных клеток, являясь основными активаторами цитолитических и фагоцитирующих эффекторов иммунитета:-ИФ является незаменимым фактором дифференцировки В-лимфоцитов.
ВИЧ-инфекция характеризуется прогрессирующей дисфункцией иммунной системы, которая сопряжена с глубокими нарушениями в системе ИФ. Для удобства изложения ВИЧ-инфекция разделена на 3 традиционные стадии: 1) ВИЧ-сероконверсию, когда в сыворотке периодически выявляются антитела к ВИЧ, но другие клинические признаки отсутствуют, 2) стадию генерализованной неспецифической лимфаденопатии и 3) клинически выраженный СПИД. Дефекты в системе ИФ прослеживаются на каждой из стадий, но их характер и глубина совершенно различны.
ВИЧ обладает интерфероногенными свойствами, обусловленными наличием в лидерной последовательности РНК 3`-LTR до промотора SP-6 двойной спирали, состоящей из 40 пар нуклеотидов. Эта особенность генома ВИЧ указывает также на потенциальную чувствительность вируса к индуцируемым ИФ классическим противовирусным механизмам - дсРНК-зависимым 2`, 5`-олиго(А)синтетазе и протеинкиназе фактора инициации синтеза белка eiF2. Однако под действием ИФ не происходит полного подавления репродукции ВИЧ. В этом отношении ретровирусы принципиально отличаются от литических вирусов. При действии ИФ активность обратной транскриптазы и синтез вирусных белков снижаются только на 70%.
Полного подавления репродукции ВИЧ удалось добиться на экспериментальной модели при постоянной продукции ИФ эндогенно. В клетки почек зеленых мартышек ввели плазмиду, содержащую ВИЧ-промотор (3`-LTR), интегрированный с геном человеческого 2-ИФ, экспрессия которого регулируется трансактивирующим фактором - продуктом гена tat. Экспрессия плазмидного гена -ИФ в трансфектных клетках приводила к активации внутриклеточной 2`, 5`-олиго(А)синтетазы. Возможность полной элиминации ВИЧ из культуры при постоянной экспрессии ИФ эндогенно представляет значительный интерес. Хотя механизм этого явления пока не установлен, все же не вызывает сомнения необходимость изучения индукторов ИФ по крайней мере на 1 стадии ВИЧ-инфекции.
В клинических наблюдениях были в основном подтверждены экспериментальные данные о неполном подавлении ВИЧ при лечении экзогенным ИФ. При ежедневном подкожном введении 35.000.000 МЕ рекомбинантного -ИФ в течение 8-12 недель больным СПИД, у них отмечено значительное, а некоторых даже полное подавление вирусных антигенов в циркуляции. Однако какого либо влияния ИФ-терапии на динамику циркулирующих антител к кору ВИЧ обнаружено не было, что подтверждает персистенцию инфекции.
У здоровых лиц ИФ в сыворотке, как правило, не обнаруживается или циркулирует в предельно низких титрах. При различных вирусных инфекциях, а также онкологических и аутоиммунных заболеваниях ИФ в сыворотках появляется постоянно, причем титр его нарастает параллельно тяжести заболевания. К настоящему времени накоплен обширный материал, позволяющий заключить, что появление ИФ в сыворотке может служить биологическим маркером вирусной инфекции, а во многих случаях и онкологического заболевания. Причем циркулирующий ИФ, как правило, относится к виду но по крайней мере, часть циркулирующего ИФ отличается необычным свойством -кислотолабильностью.
Присутствие кислотолабильного -ИФ в циркуляции у больных СПИД было установлено уже на ранних этапах исследования инфекции: он обнаруживался у 60-80% больных с генерализованной лимфоаденопатией и клинически выраженным СПИДом. Титр циркулирующего кислотолабильного -ИФ нарастал по мере увеличения клинической тяжести инфекции: на стадии генерализованной лимфаденопатии титр в среднем был равен 7,7 МЕ/мл, а при проявлении клинических симптомов СПИД - 28,7 МЕ/мл. В других наблюдения было показано, что увеличение титра циркулирующего ИФ коррелировало с гематологическими симптомами иммунодефицита. Обратная корреляция прослеживалась с общим числом лимфоцитов и с субпопуляцией Т-хелперов, и прямая - с концентрацией IgA в сыворотке.
По данным многих исследований появление циркулирующего в крови кислотолабильного -ИФ обнаруживается в среднем за 6,5 месяцев до клинических симптомов СПИДа и может служить наиболее ранним прогностическим показателем.
Однако, кислотолабильный -ИФ обнаруживался только у 65-70% больных. Остальные серопозитивные больные не имели этого диагностического признака. Этот факт до настоящего времени не получил объяснения и заслуживает специального исследования.
Таким образом, наличие циркулирующего кислотолабильного -ИФ в постоянных титрах >16МЕ/мл в группе риска является прогностическим признаком и указывает на высокую вероятность (60-80%) развития клинических симптомов СПИДа в течение нескольких месяцев.
Другой особенностью ВИЧ-инфекции, которая тоже имеет диагностическое значение, является присутствие в циркуляции ингибитора (или инактиватора ИФ).
Известно, что в физиологической норме в сыворотке периферической крови могут обнаруживаться ингибиторы ИФ типа 1 ( иПри вирусной инфекции частота их выявления существенно возрастает.
При изучении сыворотки больных СПИД осложненным саркомой Капоши, ингибитор выявлен у 80%, причем у двух третей из них его титр был достаточен для инактивации более 50 МЕ/мл -ИФ. Вместе с тем, в другой группе больных с саркомой Капоши, не имевших диагностических признаков СПИДа, ни у одного больного ингибитора в сыворотке не было. Не выявлялся ингибитор и у неосложненных больных с лимфоаденопатией. Таким образом, циркулирующий ингибитор ИФ типа 1 является диагностическим признаком, проявляющимся на III стадии ВИЧ-инфекции, одновременно с клиническими признаками СПИДа.
Химическая природа ингибитора ИФ типа 1 изучена еще недостаточно. По некоторым данным его структура напоминает структуру -ИФ.
Взаимодействие ИФ с клеткой осуществляется через специфические рецепторы с исключительно высоким аффинитетом. Иммунобиологические эффекты ИФ реализуются только через рецепторы. ИФ типа 1 имеют общий рецептор с мол. массой 110 кД, распознающая часть является ганглиозидом. Рецептор -ИФ состоит из 2-х белковых компонентов с мол. массой 50 и 90 кД.
Экспрессия рецепторов на клетке подвержена функциональным изменениям и подавляется под действием ИФ. Постоянное присутствие кислотолабильного-ИФ в циркуляции при ВИЧ-инфекции приводит к подавлению экспрессии клеточных рецепторов к ИФ типа 1, в то же время экспрессия рецепторов к ИФ типа 2 на всех клинических стадиях не изменяется.
Биологические эффекты ИФ обусловлены специфическими изменениями в метаболизме клетки. Известно, что под действием ИФ в цитоплазме появляется более 20 новых белков. Наиболее изученным ИФ-зависимым ферментом является 2`,5`-олиго(А)синтетаза, которая активируется всеми 3 видами ИФ (и в присутствии двуспиральной РНК по крайней мере из 20 нуклеотидов. Под действием этого фермента на основе АТФ синтезируется ряд коротких полиаденилатов, соединенных необычной 2`-5`-фосфодиэфирной связью. 2`,5`-олигоаденилаты являются сильными активаторами клеточных эндонуклеаз, в частности РНКазы L. Активация эндонуклеаз предотвращает считывание вирусной информации.
При ВИЧ-инфекции отмечены деформации в системе 2`,5`-олиго(А)синтетазы. У больных на стадии генерализованной лимфаденопатии активность этого фермента значительно возрастает, а при осложнении СПИД оппортунистическими инфекциями или саркомой Капоши достигает максимальных значений.
При этом повышенный фон активности 2`,5`-олиго(А)синтетазы у больных СПИДом сопровождается снижением чувствительности фермента к активирующему действию ИФ.
У больных СПИДом описан дефект и в другом звене 2`,5`-олиго(А)синтетазы - на уровне РНКазы L. Уже на стадии генерализованной лимфаденопатии активность этого фермента была в среднем на 55% ниже, чем у здоровых доноров.
Цитолиз антигенно измененных клеток естественными киллерами (ЕК) является важным механизмом противовирусного иммунитета. Детерминантами для иммунного распознавания мишеней для ЕК служат антигены главного комплекса гистосовместимости, поэтому их активность не зависит от сенсибилизации, присутствия антител или комплемента. ВИЧ-инфицированные лимфоциты оказываются высокочувствительными к действию ЕК. Моноциты и макрофаги, инфицированные ВИЧ, также интенсивно подвергаются цитолизу ЕК, выделенными из крови ВИЧ-серонегативных людей. Однако, ЕК, выделенные от ВИЧ-серопозитивных людей с давностью сероконверсии 15 месяцев, обладали уже в 2-2,5 раза меньшей активностью. Еще более глубокая депрессия ЕК отмечена у 90% больных с клинически выраженным СПИДом.
Известно, что цитолитическая функция находиться под аутокринным-паракринным контролем ИФ. При взаимодействии с антигенно измененной клеткой-мишенью ЕК продуцируют илиИФ, обеспечивая рекрутирование эффекторных клеток из предшественников и их активацию.
В то же время, ЕК, выделенные от ВИЧ-серопозитивных доноров или больных СПИДом при взаимодействии с клетками-мишенями продуцировали лишь незначительное количество ИФ. Это указывает на глубокую депрессию ЕК у больных с клинически выраженным СПИДом.
Итак, из всего вышесказанного становится ясно, что система ИФ вовлечена в патогенез ВИЧ-инфекции. Глубина деформаций системы ИФ возрастает параллельно с утяжелением клинических проявлений инфекции. На стадии сероконверсии основным дефектом, имеющим прогностическое значение, является появление в сыворотке периферической крови больного кислотолабильного -ИФ. Циркулирующий ИФ неизбежно приводит к подавлению экспрессии клеточных рецепторов к ИФ типа 1 и постоянному повышенному уровню активности 2`,5`-олиго(А)синтетазы. На стадии генерализованной лимфаденопатии указанные дефекты продолжают углубляться и начинает проявляться рефрактерность клеток крови к продукции -ИФ в ответ на вирусную или антигенную индукцию. Одновременно (а может быть и вследствие этого) снижается цитолитическая активность ЕК в отношении ВИЧ-инфицированных клеток. Снижается активность РНКазы L.
На стадии клинически выраженного СПИДа депрессия системы ИФ еще больше нарастает, в сыворотке появляется ингибитор/инактиватор -ИФ.
При анализе состояния системы ИФ при ВИЧ-инфекции необходимо учитывать, что все описанные выше явления иммунодепрессии проявляются только в отношении ИФ типа 1, главным образом -ИФ. СистемаИФ на всех стадиях ВИЧ-инфекции каким-либо существенным деформациям не подвергается.
Схема иммунологических изменений при развитии ВИЧ-инфекции.
Антитела к белку р24 Неспецифические Функция моноцитов Антитела к
Антитела к оболочке иммуноглобулины и макрофагов белку р24
вируса Специфические
Ответ на антигены, Активность иммуноглобулины
Цитотоксические или к которым имеется природных
супрессорные Т-клетки иммунологическая клеток-киллеров Клетки CD8
CD8 память Клетки CD4
Клетки
CD4/CD8<1 Клетки CD4
Заражение ПГЛ Болезни, ассоциированные СПИД
со СПИДом
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ. Клиническая картина при заражении ВИЧ может быть весьма разнообразной, начиная от острой сероконверсии и кончая полностью выраженным истинным СПИДом много лет спустя. Инфекция может протекать бессимптомно или проявляться клинически.
Начальный период после первого контакта с вирусом обычно бывает бессимптомным и может продолжаться до шести недель. Когда наконец появляются симптомы, они часто носят неспецифический характер. Возникает так называемая болезнь сероконверсии. В период острой сероконверсии наблюдается заболевание , сходное с воспалением желез: больной страдает от лихорадки, недомогания, вялости, болей в мышцах и суставах, катара горла, лимфаденопатии. В начальном периоде также может отмечаться обратимая энцефалопатия с потерей ориентации, памяти, сдвигами сознания, острый менингит, миелопатия и невропатия.
Вероятно, не у всех, у кого произошла сероконверсия, дело дойдет до хронической инфекции. У многих возможен переход инфекции в латентную фазу. Тем не менее, и в эту фазу больной является инфекционным, хотя и не в такой степени как в период сероконверсии или в стадию выраженного СПИДа.
Хроническая инфекция тоже может быть бессимптомной. Ее известные клинические признаки включают цитопению, минорные сопутствующие инфекции и кожные заболевания, лимфаденопатию, а также СПИД-ассоциированный комплекс.
Классификация состояний при инфекции ВИЧ.
Группа 1: Острая инфекция
Группа 2: Бессимптомная инфекция
Группа 3: Персистирующая генерализованная лимфаденопатия
Группа 4: Другие болезни
Подгруппа А: Конституциональные заболевания
Подгруппа В: Неврологические заболевания
Подгруппа С: Вторичные инфекционные болезни
Категория С1: Вторичные инфекционные болезни, перечисленные в списке CDC как имеющие отношение к СПИДу
Категория С2: Другие вторичные инфекционные болезни
Подгруппа D: Вторичные злокачественные опухоли
Подгруппа E: Другие заболевания
ПЕРСИСТИРУЮЩАЯ ГЕНЕРАЛИЗОВАННАЯ ЛИМФАДЕНОПАТИЯ.
Лимфаденопатию, наблюдаемую при хронической инфекции, называют синдромом персистирующей генерализованной лимфаденопатии. Определение этого синдрома таково: увеличенные лимфоузлы не менее 1 см в диаметре в двух или более несоприкасающихся внепаховых локусах, сохраняющие свой вид по меньшей мере три месяца в отсутствие какого-либо текущего заболевания или лечения, которое могло бы вызвать такой эффект.
Лимфоузлы бывают затронуты симметричны; чаще всего увеличиваются узлы, расположенные в передней и задней шейных цепочках, а также в подмышечной и подчелюстной областях, реже - в субмеатальной, заушной, эпитрохлеарной и забрюшинной областях. Примерно у трети больных имеется также ассоциированная спленомегалия. Биопсия узла обычно выявляет доброкачественную фолликулярную гиперплазию, за которой на более поздних стадиях следует инволюция фолликулов. В большинстве случаев в биопсии нет никакой необходимости. Показания к произведению биопсии будут рассмотрены далее.
СПИД-АССОЦИИРОВАННЫЙ КОМПЛЕКС.
Если у больного имеются какие-либо общие симптомы или признаки СПИДа без дополнительных инфекций или опухолей, встречающихся на поздних стадиях заболевания, то это состояние описывают как СПИД-ассоциированный комплекс. Этот термин охватывает широкий круг явлений. В настоящее время, когда разработан тест на антитела, это понятие имеет меньшее значение, однако оно все еще может быть полезным в клинической практике для выделения той группы больных, в которой можно ожидать перехода в стадию настоящего СПИДа. Более надежные клинические показатели для оценки прогноза будут рассмотрены чуть ниже.
И наконец, хроническая инфекция ВИЧ сопровождается рядом минорных добавочных инфекций и кожных заболеваний. Сюда относятся себоррейный дерматит и волосистая лейкоплакия, а также вирусные, бактериальные и грибковые инфекции, которые более подробно будут описаны далее.
Диагноз “СПИД-ассоциированный комплекс” ставиться в том случае, если пациент имеет более двух симптомов, сохраняющихся в течение трех или более месяцев, а также аномалии по данным более чем двух лабораторных тестов.
Симптомы или признаки
Лихорадка: ~ 38шС, перемежающаяся или постоянная
Потеря веса: > 10%
Лимфоузлы: персистирующая генерализованная лимфаденопатия
Диарея: перемежающаяся или постоянная
Быстрая утомляемость
Ночной пот
Отклонения в лабораторных показателях
Лимфопения, лейкопения
Тромбоцитопения
Анемия
Пониженное соотношение между CD4 и CD8
Пониженное количество Т-хелперов
Угнетенный бластогенез
Повышенный уровень гамма-глобулинов
Кожная анергия
ОППОРТУНИСТИЧЕСКИЕ ИНФЕКЦИИ И ОПУХОЛИ.
Два основных проявления СПИДа - это опухоли и ряд оппортунистических инфекций. Саркома Капоши - самый распространенный и описанный первым пример; в настоящее время выделяют и другие опухоли: неходжкиновская лимфома (обычно экстранодальная) и плоскоклеточные карциномы ротовой полости и прямой кишки. У большинства больных вначале возникает пневмония, вызванная Pneumocystis carinii, за которой следуют другие дополнительные инфекции и саркома Капоши.
Разнообразные сопутствующие СПИДу инфекции затрагивают практически любую систему организма. Больше всего распространена и характерна для больных как в США, так и в Великобритании пневмоцистная пневмония. Возбудителями пневмонии у больных СПИДом могут быть и другие организмы (см. таблицу).
У больных СПИДом в Африке саркома Капоши может сочетаться с так назывемой болезнью Слима (диарея) и другими оппортунистическими инфекциями. Более типичны для африканского континента - туберкулез, криптококкоз, криптоспоридиоз.
Спектр клинических проявлений все время расширяется. Первоначально описанные проявления саркомы Капоши и певмоцистной пневмонии относились к болезни с коротким инкубационным периодом. Однако у тех больных, у которых СПИД развивается после более длительного периода, клиническая картина может быть иной. Это как будто уже наблюдается: например, появляются новые виды опухолей и неврологических заболеваний. Наконец, накапливаются данные в пользу того, что поздняя стадия болезни сопряжена с лимфоидной интерстициальной пневмонией, гранулематозным гепатитом и энтеропатией.
Средняя продолжительность жизни больных СПИДом после постановки диагноза варьирует в зависимости от имеющихся клинических проявлений. В Америке она составляет около 9 месяцев для больных с пневмоцистной пневмонией и 31 месяц для больных с саркомой Капоши.
Благодаря многообразию проявлений инфекции ВИЧ - от бессимптомного состояния до развернутого СПИДа - Центры контроля над заболеваниями предприняли попытку разделить весь диапазон инфекций на четыре группы.
Синдром Признаки Типичные возбудители
______________________________________________________________________________
Простейшие Вирусы Бактерии Грибы
______________________________________________________________________________
Поражения Кашель, затруд- P. Carinii Цитомегаловирус Микобактерии Криптококк
легких ненность дыха- Вирус простого Str. pneumoniae
ния, лихорадка, герпеса H. influenzae
гипоксия B. catarrhalis
Видимые на рент- Стрептококки
гене инфильтраты группы В
_____________________________________________________________________________________
Поражения Менингит, Токсоплазма Цитомегаловирус Микобактерии Аспергиллы,
ЦНС энцефалит или Вирус простого криптококк,
локальные пораже- герпеса Candida
ния Паповавирусы
_____________________________________________________________________________________
Поражения Дисфагия Криптоспоридии Цитомегаловирус M. Avium Candida
кишечника Обильная диарея Isospora belli Вирус простого M. tuberculosis
Диарея с кровью Микроспоридии герпеса Сальмонелла
Колит
_____________________________________________________________________________________
Гипертермия Лихорадка Могут участвовать все возбудители очаговых или рассеянных
неизвестного Потеря веса инфекций
происхожде- Лимфаденопатия
ния
_____________________________________________________________________________________
Все упомянутые выше заболевания при их выявлении могут косвенно указывать на наличие иммунодефицитного состояния, то есть являются ВИЧ-маркерными. Ниже приводится полный список ВИЧ-маркерных заболеваний .
Протозойные гельминтозные
Криптоспоридиоз - диарея, продолжающаяся более 1 месяца
Изоспороз - то же
Пневмония, вызванная Pneumocystis carinii
Стронгилоидоз - пневмония, расстройства ЦНС, рассеянные патологии
Токсоплазмоз - пневмония, расстройства ЦНС
Грибковые
Аспергиллез - расстройства ЦНС, рассеянные патологии
Кандидоз - Поражения бронхов, легких пищевода
Криптококкоз - расстройства ЦНС, легочные и рассеянные патологии
Гистоплазмоз - рассеянные патологии
Бактериальные
Атипичный микобактериоз - рассеянные патологии, вызванные микобактериями, но не возбудителями туберкулеза или лепры.
Вирусные
Вызванные цитомегаловирусом - поражения легких, кишечника и ЦНС
Вызванные вирусом простого герпеса - тяжелые поражения кожи и слизистой, длящиеся более месяца; поражения легких, кишечника, рассеянные патологии
Прогрессирующая многоочаговая лейкоэнцефалопатия
Онкологические
Саркома Капоши - в любом возрасте
Церебральная лимфома
Неходжкинская лимфома - диффузная, недифференцированная, образована В-клетками или клетками неизвестного фенотипа
Лимфоретикулярные новообразования - по меньшей мере через 3 месяца после оппортунистических инфекций
Прочие
Хронический лимфоидный интерстициальный пульмонит у детей до 13 лет
Теперь опишем последовательность развития клинических симптомов при ВИЧ-инфекции.
Примерно через 2-4 недели после внедрения ВИЧ у половины зараженных появляется лихорадка, длящаяся от 2 до 10 дней, увеличиваются лимфатические узлы, печень и селезенка, снижается количество лимфоцитов в крови. Затем все проходит будто бы бесследно: инфицированный человек ни на что не жалуется. Однако через несколько месяцев (а чаще лет) у него медленно но неуклонно начинают обнаруживаться симптомы заболевания.
Отмечено, что при переливании крови, инфицированной ВИЧ, средняя продолжительность инкубационного периода составляет у детей два года, у пожилых лиц - пять лет, у людей среднего возраста - восемь.
Обычно, прежде чем развернуться полной картине заболевания, у больного наступает пре-СПИД: постепенно повышается температура до 38-39шС, отмечается обильное потоотделение, особенно в ночное время, резкая утомляемость, разбитость, исчезает аппетит. Важный, а то и самый ранний признак - стойкое увеличение лимфатических узлов: шейных, подчелюстных, затылочных, но, как правило, не паховых. Расстраивается деятельность кишечника: частый водянистый стул. Происходит прогрессирующая потеря веса.
Все эти симптомы рассматриваются как пре-СПИД только в том случае, если у больных установлено вирусоносительство и если при обследовании иммунной системы обнаруживается снижение количества Т-хелперов при относительно увеличенном или неизмененном количестве Т-супрессоров. В присутствии вируса особое значение имеет лимфаденопатия неясной этиологии. И чтобы исключить лимфомы, сифилис, туберкулез, делается биопсия лимфатических узлов.
В дальнейшем болезненные симптомы начинают нарастать. Похудение может достигать 10-15 килограммов и более. Нередко присоединяются оппортунистические инфекции, и прежде всего пневмоцистная пневмония. У значительной части больных развивается саркома Капоши. Наличие ее у лиц моложе 60 лет в сочетании с положительными результатами лабораторных исследований (появление антител к ВИЧ, снижение числа Т-хелперов) служит несомненным доказательством СПИДа. Так же как лимфомы головного мозга.
Злокачественные опухоли характерны для 40% больных СПИДом, причем из этой цифры 85% приходится на саркому Капоши и 10% на злокачественные лимфомы.
Заболевание длится от нескольких месяцев до 4-5 лет. Исход его - смерть.
При СПИДе могут быть периодические обострения и временные улучшения. У разных больных отмечается преобладание тех или иных симптомов: у одних поражаются преимущественно легкие, у других - нервная система, третьих мучает острая диарея и др. Но, как правило, самый ранний признак - лимфаденопатия, тем более если она продолжается больше двух месяцев без видимых причин.
При диагностике принимаются во внимание эпидемиологические данные: особенно подозрительно, если саркома Капоши и лимфомы диагностируются у гомосексуалистов, наркоманов, лиц с беспорядочными половыми связями.
В последние годы вызывают тревогу сопровождающие СПИД, а также пре-СПИД признаки поражения центральной нервной системы. Наиболее опасный и частый симптом - прогрессирующее слабоумие (деменция) как результат атрофии коры головного мозга. Этот симптом регистрируется сейчас примерно у 50% больных. По мнению многих специалистов, мозговые нарушения, преимущественно слабоумие, могут развиться у каждого человека, инфицированного ВИЧ. При вскрытии у 2/3 больных, умерших от СПИДа, наблюдалась атрофия ткани мозга.
Предполагается, что у взрослых нарушения нервной деятельности могут наступить даже через 20-30 лет после заражения и что средняя продолжительность скрытого периода “зреющего” слабоумия составляет 15 лет. Отмечаются также поражения сосудов мозга, менингит. Больные часто жалуются на головную боль, снижение остроты зрения. Возникают абсцессы, вызванные токсоплазмами, микобактериями, грибами Candida.
Размножение ВИЧ в клетках мозга приводит к утрате кратковременной памяти, нарушению координации, мышечной слабости, расстройству речи, психики. При этом поражение мозга не всегда сопровождается иммунодефицитом.
При тщательном неврологическом обследовании те или иные “сбои” в работе мозговой деятельности - ослабленное внимание, затрудненная координация движений, замедленность реакций - удается распознать и у так называемых бессимптомных носителей. Поэтому чрезвычайно важно подвергать такому обследованию всех инфицированных лиц, тем более если от них зависит жизнь и здоровье других людей. В случае, если у них подтверждаются соответствующие изменения в функциях мозга, их отстраняют от работы.
Врачей очень тревожит, что эпидемия слабоумия распространится среди молодежи, инфицированной ВИЧ.
Вместе с тем самые ранние и выраженные мозговые нарушения наблюдаются у детей, особенно - у заразившихся в утробе матери или сразу после рождения.
Особенности инфицирования и заболевания детей.
ВИЧ-инфекция у детей может быть связана с заболеванием матерей, переливанием зараженной крови при гемофилии, некачественными инъекциями, наркоманией. Перенос ВИЧ от зараженных матерей наблюдается у 25-30% потомков.
Сама по себе беременность благоприятствует развитию СПИДа, так как она обычно сопровождается иммунодепрессией, особенно депрессией клеточного иммунитета. При нормальной беременности, главным образом в третьем триместре, отношение числа Т-хелперов к Т-супрессорам снижается. Иммунитет восстанавливается примерно через 3 месяца после родов. Имеются указания на повышенный риск беременных женщин в отношении СПИДа, поскольку роды у инфицированных ВИЧ, очевидно, способствуют более быстрому развитию заболевания.
Заражение детей происходит преимущественно в матке, родовых путях и после родов. ВИЧ способен проникать через плаценту. Заражение потомков может быть связано и с грудным вскармливанием, так как вирус выделен из молока инфицированных матерей.
Описаны случаи, когда инфицированные матери рожали близнецов, из которых только один был инфицирован.
Дети зараженные от матерей начинают болеть через 4-6 месяцев после инфицирования, и большинство из них обычно погибает в течение 2 лет.
СПИД у детей характеризуется рядом особенностей. У них меньше, чем у взрослых, продолжительность скрытого периода, часто он длится не годы, а месяцы. Диагностировать заболевание у детей труднее, особенно в первый год жизни. Объясняется это прежде всего тем, что определение у них антител к ВИЧ связано со значительными сложностями: неясно, получил ли больной антитела из крови матери через плаценту или же они образуются у него в результате заражения. Проблему можно решить, выделив вирус. Но и это сложно. Тут нередко наблюдается рецидивирующая инфекция дыхательных путей - лимфоидная интерстициальная пневмония.
Поражение ЦНС у детей может выражаться в утрате основных вех развития. Например, ребенок, который в соответствии с возрастом мог уже сидеть и говорить, теряет эту способность. У него перестает увеличиваться размер головы. К начальным признакам заболевания относится также отсутствие прибавки в весе, хронический понос, бактериальные инфекции.
Наиболее важная иммунологическая особенность детей, больных СПИДом, - наличие в их крови исключительно высокого содержания иммуноглобулинов и одновременно неспособность вырабатывать антитела при введении антигенов, которые вызывают в нормальных условиях образование антител, в частности к ВИЧ.
Таким образом, если в период новорожденности в результате заражения через плаценту или при переливании крови происходит заражение ВИЧ, то прогноз для этих детей исключительно неблагоприятный: у них следует ожидать прогрессирующего развития заболевания особенно с поражением ЦНС, обусловленного непосредственно ВИЧ.
Все сказанное выше подводит к однозначному выводу: детей не должны рожать не только инфицированные женщины, но и те, что общаются с инфицированными партнерами.
ВИРУС ИММУНОДЕФИЦИТА ЧЕЛОВЕКА ТИПА 2 (ВИЧ-2).
ВИЧ-2 был впервые выделен в 1985 году от больных СПИДом в Гвинее-Биссау и Островах Зеленого Мыса. В дальнейшем были изучены особенности возбудителя и в меньшей степени клинико-эпидемиологические особенности инфекции. В настоящее время доказано, что ВИЧ-2 и ВИЧ-1 являются самостоятельными инфекциями, поскольку имеются различия в особенностях возбудителей, клинике и эпидемиологии. Ниже приводится обзор сведений, полученных к настоящему времени по ВИЧ-2.
Географическое распространение. Важной отличительной особенностью инфекции ВИЧ-2 является ее преимущественное распространение в странах Западной Африки. Антитела к ВИЧ-2 обнаружены у жителей 15 африканских стран: Анголы, Буркина-Фасо, Ганы, Гамбии, Гвинеи, Гвинеи-Биссау, Зимбабве, Камеруна, Кот Д`Ивуар, Мали, Мозамбика, Сенегала, Сьерра-Леоне, Того и Центральноафриканской Республики. Инфекция за пределами Западной Африки регистрируется довольно редко.
Следует иметь в виду, что в некоторых странах Западной Африки, таких как Гвинея-Биссау, ВИЧ-2 является практически единственным циркулирующим среди населения вирусом иммунодефицита. В Сенегале, Гамбии, Мали и Мозамбике в общей массе инфицированных ВИЧ на долю инфицированных ВИЧ-2 приходится от 37 до 68%. Обращает на себя внимание наличие в этих странах значительного числа лиц (до 50%) с двойной инфекцией.
Но инфицированные ВИЧ-2 крайне редко встречаются в Европе и Америке. В связи с этим встает вопрос о целесообразности обследования в этих странах доноров на инфицированность ВИЧ-2. Обследование с двумя тест-системами значительно дороже. Однако чтобы не проглядеть даже единичных случаев, предлагается использовать методы, которые наиболее часто дают перекрестные реакции между ВИЧ-1 и ВИЧ-2. Либо предлагается разработать такие тест-системы, которые содержат одновременно антигены ВИЧ-2 и ВИЧ-1. В частности такие системы (combitest) уже предложены рядом коммерческих фирм. Поскольку риск заражения ВИЧ-2 в Европе и Америке весьма мал, в случаях недостаточности комбинированных тест-систем рекомендуется обследовать на ВИЧ-2 только тех доноров, которые имели связи с Западной Африкой или половые контакты с выходцами из этих стран. Рекомендуется также обследовать на ВИЧ-2 тех доноров, у которых неопределенная реакция на ВИЧ-1.
Строение вируса. Проведенные детальные исследования свойств ВИЧ-2 показали, что при определенном сходстве с ВИЧ-1, он отличается от последнего по антигенной структуре и по последовательности оснований в нуклеиновых кислотах. ВИЧ-2 более близок по своим свойствам (в том числе по антигенной структуре и составу генетического материала) к вирусу иммунодефицита обезьян (SIV/ВИО), чем к ВИЧ-1.
ВИЧ-1 и ВИЧ-2 инфицируют те же популяции клеток, связываются с теми же CD4-рецепторами. Проведен анализ полного генома ВИЧ-2. В целом его организация сходна с таковой ВИЧ-1. В дополнение к классическим ретровирусным генам gag, pol и env оба вируса содержат гены vif, vpr, tat, nef и rev.
Одним из наиболее существенных различий между вирусами является наличие у ВИЧ-1 гена vpu, не обнаруживаемого у ВИЧ-2. В тоже время ВИЧ-2 содержит ген vpx, которого нет у ВИЧ-1. Продуктом гена vpu является белок с молекулярной массой 14-16 кД, а гена vpx - белок с молекулярной массой 12-14 кД. Продукт гена vpu весьма важен для созревания и высвобождения вируса из клетки. Гены tat, ref и nef необходимы для вирусной репликации и экспресси вирусного генома. Роль генов vif и vpr пока неясна. Полагают, что vif может играть роль в посттрансляционной модификации вирусных белков.
Наиболее консервативными участками геномов ВИЧ-1 и ВИЧ-2 являются гены gag и pol. Они соответственно имеют 57 и 59% гомологии. Менее консервативен участок, кодирующий оболочечные белки, особенно содержащиеся в наружной части оболочки вируса (37% идентичных аминокислот). Было установлено, что большинство цистеиновых остатков в ВИЧ-1 и ВИЧ-2 стабильно, это свидетельствует о том, что третичная структура белка может иметь важное функциональное значение. Трансмембранная часть оболочечного белка - gp41- более стабильна, чем наружная - gp120. Важную роль в цитопатогенности ВИЧ отводят С-концу трансмембранного белка. N-терминальный конец этого белка является уникальным регионом, специфичным для каждого из ВИЧ. На основании этой уникальности можно осуществить дифференциацию ВИЧ-1 и ВИЧ-2. ВИЧ-2 отличается от ВИЧ-1 по оболочечным гликопротеинам и в меньшей степени - по сердцевинным антигенам. Сыворотки большинства людей, инфицированных ВИЧ-2, не реагируют с оболочечными антигенами ВИЧ-1, но дают перекрестные реакции с антигенами, кодируемыми геном gag.
Имеются различия в молекулярной массе белков ВИЧ-1 и ВИЧ-2. В частности, наружный оболочечный гликопротеин имеет молекулярную массу 120 кД, у ВИЧ-2 этот белок имеет мол. Массу 140 кД. Трансмембранный белок у ВИЧ-2 - 36 кД, а у ВИЧ-1 - 41кД Белки кодируемые геном gag у ВИЧ-1 имеют мол. Массу 24 и 17 кД, а у ВИЧ-2 - 26 и 15 кД.
Эпидемиология. Эпидемиология ВИЧ-2 изучена относительно мало. Полагают, что распространение ВИЧ-2 еще только начинается. ВИЧ-2 передается теми же путями, что и ВИЧ-1. Вирус выделен как от лиц с клинически выраженным СПИДом, так и от бессимптомных носителей. Поскольку инфицированные лица не являлись ни гомосексуалистами, ни наркоманами, полагают, что ВИЧ-2 передается преимущественно путем гетеросексуальных половых связей. При обследовании различных групп населения было выявлено, что число сероположительных результатов к ВИЧ-2 было в 10-15 раз больше среди проституток, чем среди других групп населения. В странах Западной Африки число сероположительных с антигенами ВИЧ-1 лиц было значительно ниже, чем с таковыми ВИЧ-2. Вместе с тем ВИЧ-2 практически отсутствовал в центральноафриканских странах. Трехлетнее наблюдение за проститутками показало, что болезнь у зараженных ВИЧ-2 развивается значительно медленнее, чем при инфицировании ВИЧ-1. Через 3 года после выявления серопозитивности к ВИЧ-2 не было выраженных лимфаденопатией или СПИДа, что, возможно, свидетельствует о меньшей вирулентности ВИЧ-2. Исходя из различий в цитопатической активности различных штаммов ВИЧ-2, полагают, что штаммы ВИЧ-2 могут широко варьировать в патогенности и вирулентности для человека. Вич-2 менее вирулентен, чем ВИЧ-1. Обращает на себя внимание, что процент инфицированности в группах повышенного риска увеличивается с возрастом. В частности, на юге Сенегала в группе из 140 проституток инфицированность ВИЧ-2 достигала 40%. Наибольшее число положительных результатов было выявлено у женщин старше 50 лет, имевших наибольшее число половых партнеров.
Систематические наблюдения свидетельствуют о том, что ВИЧ-2 не передается столь легко, как ВИЧ-1. Для передачи ВИЧ-2 необходимы повторные контакты. Основным путем передачи инфекции является гетеросексуальный путь (в 40% семей были сероположительны оба супруга). Не зарегистрированы случаи вертикальной передачи ВИЧ-2, не наблюдалась также передача инфекции вследствие кормления грудью. Было обнаружено, что большинство серопозитивных детей получило инфекцию при переливании крови. Однако недавно сообщено о случае вертикальной передачи от матери к ребенку. Предварительные данные свидетельствуют, что инкубационный период является довольно длительным.
Судя по серологическим данным и по одновременному выделению обоих вирусов от одних и тех же лиц или по другим вирусным маркерам, в районах циркуляции обоих вирусов имеют место двойные инфекции. Вместе с тем, следует отметить, что случаи двойной инфекции, основанные исключительно на серологических данных, должны интерпретироваться весьма осторожно в связи с наличием перекрестных реакций.
Клинические отличия. Большинство исследователей, изучавших ВИЧ-2, нашли, что эта инфекция развивается медленнее, чем ВИЧ-1. Симптоматика СПИД вызванного ВИЧ-2 в целом не отличается от симптоматики СПИДа, вызванного ВИЧ-1. Вирус типа 2 вызывает те же клинические синдромы. У больных наблюдается резкое снижение количества циркулирующих CD4-лимфоцитов. Предварительные данные свидетельствуют о более благоприятном течении СПИДа, вызванного ВИЧ-2. У лиц инфицированных ВИЧ-2 даже через 3 года не было выраженных лимфаденопатией или СПИДа. В связи с подобными наблюдениями полагают, что риск развития СПИДа у людей, инфицированных ВИЧ-2 ниже, чем при заражении ВИЧ-1. Вместе с тем в наблюдениях Clavel 7 из 17 больных СПИДом, вызванным ВИЧ-2, умерли в течение 1 года.
Следовательно, вопрос о тяжести течения инфекции нуждается в дальнейшем изучении. Остается также неясным влияние двойной инфекции (ВИЧ-1 и ВИЧ-2) на тяжесть клинического течения.
Лабораторная диагностика. Для выделения ВИЧ-2 используют те же методы, что и для ВИЧ-1. Лимфоциты больных сокультивируются с нормальными, стимулированными ФГА лимфоцитами, при постоянном присутствии интерлейкина-2. При наличии ВИЧ-2 обнаруживаются многоядерные гигантские клетки, уменьшается общее количество клеток, появляется обратнотранскриптазная активность, которая достигает пика через 2-3 недели.
Наблюдаются широкие штаммовые вариации в скорости наступления и выраженности описанных явлений.
Что касается выявления антител к ВИЧ-2, то в целом существующие лизатные диагностические системы ИФА, предназначенные для диагностики ВИЧ-1, позволяют также распознать инфекцию ВИЧ-2 в 82-84% случаев, поскольку антигены, кодируемые генами gag и pol, обнаруживают значительную гомологию. Перекрестные реакции могут наблюдаться не только в ИФА, но и в иммуноблоте, и в реакции нейтрализации вирусов. Значительно реже перекрестные реакции имеют место при использовании конкурентного ИФА.
Вместе с тем ни один из упомянутых методов при использовании антигенов ВИЧ-1 не дает 100% чувствительности с антителами к ВИЧ-2. Это несовпадение особо отчетливо проявляется у больных СПИДом, которые по мере развития заболевания утрачивают антитела именно к антигенам, дающим перекрестные реакции. Поэтому для получения тест-систем со 100% специфичностью и чувствительностью по отношению к ВИЧ-2 рекомендуется использовать типоспецифические пептиды ВИЧ-2 полученные синтетическим или рекомбинантным методом. При смешении пептидов ВИЧ-1 и ВИЧ-2 удается в ИФА одновременно выявлять антитела к обоим вирусам (так называемый комбитест фирмы “Аббот”). Последующее испытание с раздельными антигенами позволяет провести типоспецифическую диагностику.
Большие надежды возлагаются на возможность дифференциации антител при помощи антигенов, кодируемых генами vpu и vpx, которые соответственно уникальны для ВИЧ-1 и ВИЧ-2.
В целом в группах населения, где возможна циркуляция обоих вирусов, рекомендуется ИФА, основанный на применении синтетических или рекомбинантных пептидов.
ДИАГНОСТИКА ВИЧ-ИНФЕКЦИИ.
Антитела к ВИЧ появляются начиная от трех недель до трех месяцев после инфицирования вирусом, и в дальнейшем их можно почти всегда обнаружить, даже если вирус подавляет в какой-то мере функцию лимфоцитов и выработку антител. Однако титр выявляемых нейтрализующих антител низок, а действие незначительно - они не приостанавливают заметным образом развитие инфекции и заболевания.
Для целей диагностики ВИЧ можно выделять в больших количествах из клеточных линий, очищать и использовать как антиген в серологических тестах. Существуют несколько типов тестов на антиВИЧ-АТ. Вот три из них, которые поступают в продажу:
Непрямой метод ELISA:
Антиген - проба - анти-Ig фермент
Конкурентный метод ELISA:
Антиген - проба
антиВИЧ-АТ
“Сэндвичевый” метод ELISA или агглютинация:
Антиген - проба - Антиген (с присоединенным ферментом)
ELISA - Enzyme-Linked Immunosorbent Assay или иммуноферментный анализ - ИФА
В большинстве тестов применяется коньюгат антигена с ферментом, а сигналом служит цветная реакция между специфически связанным ферментом и его субстратом. В других тестах используют радиоизотопы, антиген-флуоресцин или агглютинацию покрытых вирусом частиц латекса или желатины.
С тех пор, как в 1985 году тесты на антиВИЧ-АТ поступили в продажу, они нашли широкое применение в развитых странах и лабораториях по диагностике и переливанию крови. Точность тестов - как их чувствительность, так и специфичность - неуклонно повышается: случаи ложнонегативных и ложнопозитивных ответов становятся все реже.
В дополнение к тестам, выявляющим антитела к ВИЧ “суммарно”, существуют более тонкие тесты для выявления тех или иных компонентов иммунного ответа. Реакция на индивидуальные белки ВИЧ детально изучена методами иммуноблотинга и радиоиммунопреципитации. Наряду с этим можно определять отдельные классы иммуноглобулинов в крови и других жидкостях. Особый интерес представляют с этой позиции антиВИЧ-иммуноглобулины класса М, так как они в начале инфекции появляются раньше, чем IgG.
Для массовых исследований на антиВИЧ-АТ в неоптимальных лабораторных условиях разрабатываются упрощенные варианты тестов. Они удобны тогда, когда результат необходимо получить срочно. Рассматривается возможность использования слюны как материала для диагностики.
Во многих странах введены дополнительные процедуры, обеспечивающие достоверность результатов. Проверка состоит в том, что в случае положительного ответа проба на антиВИЧ-АТ повторяется, тестируемые материалы проверяются другими, методически независимыми способами. Хотя это и приводит к некоторой задержке, зато результат получается более точным. Наибольший остаточный риск после этого связан с тем, что у отдельных инфицированных лиц может не быть антиВИЧ-АТ, либо их концентрация будет так низка, что даже очень чувствительные тесты не смогут их выявить. Это весьма возможно на ранних стадиях инфекции, и без дальнейшего наблюдения такие случаи легко пропустить.
Помимо антител, в сыворотке присутствуют вирусные антигены, в частности, основной белок сердцевины р24. Его можно выявлять пока он еще в избытке по отношению к антителам против него, - обычно в самом начале инфекции. Тесты на ВИЧ-антиген поступили в продажу и могут использоваться в качестве дополнения к тесту на антитела. Они помогают в диагностике ранней фазы инфекции, а также в распознавании инфекции у детей. На более поздних стадиях присутствие антигенов ВИЧ в сыворотке говорит об иммунном истощении и может служить показанием для противовирусной терапии.
Виремия, на которую указывает возможность выделить ВИЧ из лимфоцитов, может быть обнаружена даже на фоне высоких титров антител к р24 и другим белкам вируса. Однако выделение вируса - процедура, требующая времени, и для успешной лабораторной диагностики ВИЧ у людей с малым количеством антител или вовсе без них важнее регулярно получать пробы в последующий период. Наблюдения за развитием инфекции с момента заражения показывают, что как титр, так и набор антител к ВИЧ обычно увеличиваются. У лиц, зараженных несколько месяцев назад или раньше почти всегда выявляется сильный антивирусный ответ.
Помимо указанных выше методов диагностики ВИЧ-инфекции в практике используют полимеразную цепную реакцию (ПЦР) и ДНК-зондовые методы. ПЦР используют для количественной оценки активности репликации вируса по числу вирусных РНК. ДНК-зондовые методы позволяют выявлять ВИЧ в форме провируса. Это особенно необходимо, когда тесты на антитела к ВИЧ и антигены дают отрицательные результаты., в латентный период болезни.
ЛЕЧЕНИЕ.
В разных странах ведутся интенсивные исследования с целью обнаружить эффективные лечебные средства против СПИДа. Основные стратегические направления этих работ - поиск противовирусных препаратов, влияющих на ВИЧ в разные стадии его размножения, и применение методов восстановления нарушенных при СПИДе функций иммунитета. Кроме того, изучаются оптимальные условия лечения развивающихся при СПИДе инфекционных осложнений и опухолей, особенно саркомы Капоши.
Один из перспективных путей - изыскание препаратов, блокирующих синтез обратной транскриптазы, то есть процесс, когда происходит переписывание генетической информации с РНК на ДНК. Сейчас наиболее перспективными в смысле противодействия обратной транскриптазе считаются аналоги нуклеотидов. Одним из первых более или менее эффективных препаратов стал азидотимидин (зидовудин, AZT). Он увеличивает среднее время выживания больных в далеко зашедшей стадии СПИДа примерно на год. Однако азидотимидин в значительной степени токсичен - от него больше других страдает костный мозг, что приводит к анемии. В настоящее время пытаются применять другие аналоги нуклеотидов самостоятельно или в различных комбинациях друг с другом. Большинство препаратов еще находиться в стадии клинических испытаний. В связи с этим клиники и лаборатории постоянно производят набор волонтеров для этого. Вот перечень основных препаратов, которые на май 1997 года находились в стадии клинических испытаний и давали наиболее эффективные результаты:
Adefovirdipivoxil - отличается ацикличным остатком сахара и одной фосфатной группой, в связи с чем легче фосфорилируется в клетке, чем другие аналоги нуклеотидов.
Beta-fluoro-ddA - флюороаналог диданозина; благодаря атому фтора усваивается лучше, чем диданозин. Эффективен против тех штаммов вируса, которые имеют множественную устойчивость к дидеоксинуклеотидам из-за мутации в 151 триплете гена обратной транскриптазы.
Didanozine - менее токсичный чем AZT, аналог нуклеотида; лучше метаболизирует в клетке.
Другое направление антивирусных препаратов - вещества, ингибирующие протеазу ВИЧ; тем самым подавляется процесс созревания вирусных белков, формируются неполноценные, неинфекционные вирусные частицы. Эти препараты также находятся в стадии клинических испытаний. Вот наиболее эффективные из них:
Ritonavir, ABT-378 (в 10 раз сильнее Ritonavir), Saquinavir, Indinavir. Эффективно комплексное использование этих препаратов.
Обнадеживающие результаты получены при применении Ro 24-7429. Его действие основано на связывании белка - продукта гена tat, то есть подавляется эффект трансактивации вируса.
Итак, эффективные противовирусные препараты получить возможно, но дело осложняет тот факт, что ВИЧ обладает очень большой изменчивостью. В одном организме можно постоянно выделять новые сероварианты вируса. В связи с этим ВИЧ быстро приобретает лекарственную устойчивость, и эффективные ранее препараты становятся почти бесполезны.
Разрабатываются и методы симптоматической и патогенетической терапии. Одна из обнадеживающих разработок - препарат из растворимых молекул CD4, способных связываться с ВИЧ и препятствовать его адсорбции на мембранах специфических клеток.
Пока же главные меры обращены на терапию саркомы Капоши, оппортунистических инфекций. Сильнее поддаются лечению заболевания, вызванные грибками, токсоплазмами. Воздействие на возбудителя пневмоцистоза не так эффективно. Еще менее действенно лечение инфекций, вызванных некоторыми вирусами, активирующимися при СПИДе.
Попытки восстановления иммунной системы с помощью ИЛ-2, интерферонов, тимусных факторов или трансплантации косного мозга не имеют значительного успеха. Наблюдаются лишь временные улучшения. При этом, иммунотерапия может спровоцировать усиление репликации ВИЧ, так как вирус размножается в основном в делящихся клетках.
ВОЗМОЖНОСТИ РАЗРАБОТКИ ВАКЦИН.
Иммунизация против болезнетворного агента, повреждающего важный компонент иммунной системы, связана с особыми трудностями. К тому же мешает сильная изменчивость вируса. Она обусловлена преимущественно накоплением мутаций. Нельзя исключить роль и генетических рекомбинаций - обмена генами между разными вариантами ВИЧ и других вирусов, которые часто находятся в организме пораженном СПИДом, а также между генами ВИЧ и клеточными генами больного. До сих пор во всех попытках иммунизации против вируса использовался очищенный или клонированный гликопротеин оболочки. У экспериментальных животных он действительно вызывает образование нейтрализующих антител к вирусу, но только к тому штамму, который был использован для иммунизации. Иногда вырабатываются нейтрализующие антитела, которые действуют на несколько штаммов, но их титр, как правило, очень низок. Более того, до сих пор точно неизвестно против какого компонента вируса направлены нейтрализующие антитела. Тем не менее оболочка вируса сохраняет свою привлекательность в качестве антигена для иммунизации, так как процесс связывания с молекулой CD4 оказался общим для всех изученных на сегодняшний день штаммов, и это говорит о возможности наличия общих эпитопов у их оболочек. Вероятно, нейтрализующие антитела к этим консервативным участкам можно получить, используя в качестве антигена антитела к CD4 (антиидиотипический метод).
Результаты экспериментов с животными позволяют думать, что важно не только то, какой из компонентов вируса используется для вакцинации, но и то, каким способом вакцина “предлагается” иммунной системе. Было показано, что в качестве вакцины могут быть весьма эффективны вирусные антигены, включенные в “искомы” - иммуностимулирующие комплексы.
Кроме того, адекватная оценка вакцин затруднена, так как пока не известен ни один вид, кроме человека, у которого ВИЧ вызывал бы подобные СПИДу заболевания (хотя у некоторых приматов возможна кратковременная инфекция).
Поэтому эффективность вакцин можно исследовать только на добровольцах. Подобные испытания уже проводятся в некоторых странах. Однако как долго придется ждать результатов изучения эффективности вакцины, если скрытый период при СПИДе длится многие годы? В этом состоит лишь одна из трудностей.
И все же некоторые перспективы уже наметились. Изучаются генноинженерные методы создания вакцины против ВИЧ: в генетический аппарат вируса коровьей оспы встраивается ген одного из белков ВИЧ. Интерес представляет работа, ведущаяся в Институте иммунологии Министерства здравоохранения России. Метод основан на применении синтетических иммуногенов, позволяющих стимулировать В-лимфоциты, минуя Т-клеточный контроль.
В мае 1997 года в США появилась публикация о том, что в ближайшее время должны начаться клинические испытания вакцины, которая действует на иммуногенные клетки слизистой оболочки половых путей. В сообщении говориться о том, что еще очень плохо изучен иммунитет связанный со слизистыми оболочками, а это могло бы очень способствовать созданию вакцины против возбудителей, передача которых связана со слизистыми оболочками.
Иммунологи предъявляют жесткие требования к вакцине: она не должна пропускать ВИЧ в клетки-мишени; нейтрализовать ВИЧ до проникновения его в мозг; обеспечивать распознавание иммунной системой всех возможных вариантов ВИЧ и гарантировать защиту всех вакцинированных.
Так обстоят дела с получением вакцины против ВИЧ. В исследования включились крупнейшие ученые ряда стран. Несмотря на чрезвычайную сложность проблемы, наметились различные пути ее решения, постоянно генерируются новые идеи, в некоторых лабораториях уже начаты клинические испытания вакцины на людях. Все это дает основания надеяться, что вакцина против вируса СПИДа будет получена. Но все же подавляющее большинство ученых, работающих в этой области, считают, что широкого применения вакцины надо ожидать не ранее, чем через 5-7 лет.
А если в ближайшее время рассчитывать на вакцину не приходится, то хотя бы для приостановки роста эпидемии необходимо перекрыть основные пути передачи ВИЧ. В связи с этим ВОЗ создала специальные, направленные на это программы. Основные пункты этих программ:
* Санитарное просвещение
* Предотвращение передачи вируса половым путем
* Предотвращение передачи вируса при инъекции наркотиков
* Предотвращение вертикальной передачи вируса
* Борьба с передачей вируса через кровь, препараты крови, донорские органы
* Профессиональные работники здравоохранения как пропагандисты
* Эпидемиологический надзор
ЗАКЛЮЧЕНИЕ.
Итак, сейчас уже многим ясно, что СПИД - одна из важнейших и трагических проблем, возникших перед всем человечеством в конце ХХ века. И дело не только в том, что в мире уже зарегистрированы многие миллионы инфицированных ВИЧ и более 200 тысяч уже погибло, что каждые пять минут на земном шаре происходит заражение одного человека. СПИД - это сложнейшая научная проблема. До сих пор неизвестны даже теоретические подходы к решению такой задачи, как очистка генетического аппарата клеток от чужеродной (в частности, вирусной) информации. Без решения этой проблемы не будет полной победы над СПИДом. А таких научных вопросов это заболевание поставило много...
СПИД - это тяжелейшая экономическая проблема. Содержание и лечение больных и инфицированных, разработка и производство диагностических и лечебных препаратов, проведение фундаментальных научных исследований и т. Д. Уже сейчас стоят миллиарды долларов. Весьма непроста и проблема защиты прав больных СПИДом и инфицированных, их детей, родных и близких. Трудно решать и психосоциальные вопросы, возникшие в связи с этим заболеванием.
СПИД - это не только проблема врачей и работников здравоохранения, но и ученых многих специальностей, государственных деятелей и экономистов, юристов и социологов.
Все должны знать и понимать, что СПИД войдет с нами в XXI век. И для того, чтобы выжить, уменьшить риск заражения и распространения заболевания, как можно больше людей во всех странах должны многое знать о СПИДе - о развитии и клинических проявлениях заболевания, его профилактике и лечении, уходу за больными, психотерапии больных и инфицированных.
Список использованной литературы.
А.С. Шевелев, СПИД - загадка века; М.,1991г.
Б.И. Веркин, Ю.Л. Волянский, Л.М. Марчук и др., Синдром приобретенного иммунодефицита. Возможные механизмы взаимодействия вируса иммунодефицита человека с клетками организма; Харьков, 1988.
М. Адлер, Азбука СПИДа; М.: Мир, 1991.
Н.К. Шарова, А.Г. Букринская, Особенности взаимодействия белков в составе вирионов ВИЧ-1; Вопросы иммунологии, 1990, т.3, №3, с. 202-206.
“Возможно выделен новый тип вируса иммунодефицита человека - ВИЧ-3”; Вопросы вирусологии, 1990, т.35, №1, с. 82.
Л.А. Кожемякин, В.Г. Бондаренко; Нестабильность генома и СПИД. Биохимия, 1992, т. 57, в. 9, с. 1417-1426.
В.П. Кузнецов. Система интерферона при ВИЧ-инфекции. Вопросы вирусологии. 1991, т. 36, №2, с. 92-96.
Т.А. Бектимиров. Вирус иммунного дефицита человека типа 2. Вопросы вирусологии. 1990, т. 35, №3, с. 180-183.
М.И. Букринский. Строение генома и экспрессия генов вируса иммунодефицита человека (обзор иностранной литературы). Вопросы вирусологии. 1987, т.32, № 6, с. 649-656.
MED/96036380. Haubrich RH, Flexner C, Lederman MM, Hirsch M, Pettinelli CP, Ginsberg R, Lietman P, Hamzeh FM, Spector SA, Richman DD. A randomized trial of the activity and safety of Ro 24-7429 (Tat antagonist) versus nucleoside for human immunodeficiency virus infection. The AIDS Clinical Trials Group 213 Team. J Infect Dis 1995 Nov;172(5):1246-52.
MED/96217734. Jacobsen H, Hanggi M, Ott M, Duncan IB, Owen S, Andreoni M, Vella S, Mous J. In vivo resistance to a human immunodeficiency virus type 1 proteinase inhibitor: mutations, kinetics, and frequencies. J Infect Dis. 1996 Jun;173(6):1379-87.
MED/96147316. Goldman AI, Carlin BP, Crane LR, Launer C, Korvick JA, Deyton L, Abrams DI. Response of CD4 lymphocytes and clinical consequences of treatment using ddI or ddC in patients with advanced HIV infection. J Acquir Immune Defic Syndr Hum Retrovirol. 1996 Feb 1;11(2):161-9.
CDC AIDS Daily Summary 5/12/97. "AIDS Vaccine Proving to Be an Elusive Goal"
Los Angeles Times--Washington Edition (05/12/97) P. A4; Cimons, Marlene.
Information presented at the 4th Conference on Retroviruses and Opportunistic Infections in Washington, DC, January 22-26, 1997.
Все иностранные статьи, статистика и некоторые другие данные по разным разделам взяты из Интернета по адресам:
gopher://gopher.niaid.nih.gov (сервер Национального института здоровья США)
http://www.niaid.nih.gov (сервер Национального института здоровья США)
http://www.aegis.gov
gopher://gopher.hivnet.org
http://noah.cuny.edu/aids
http://galen.library.uesf.edu/aids
Похожие работы


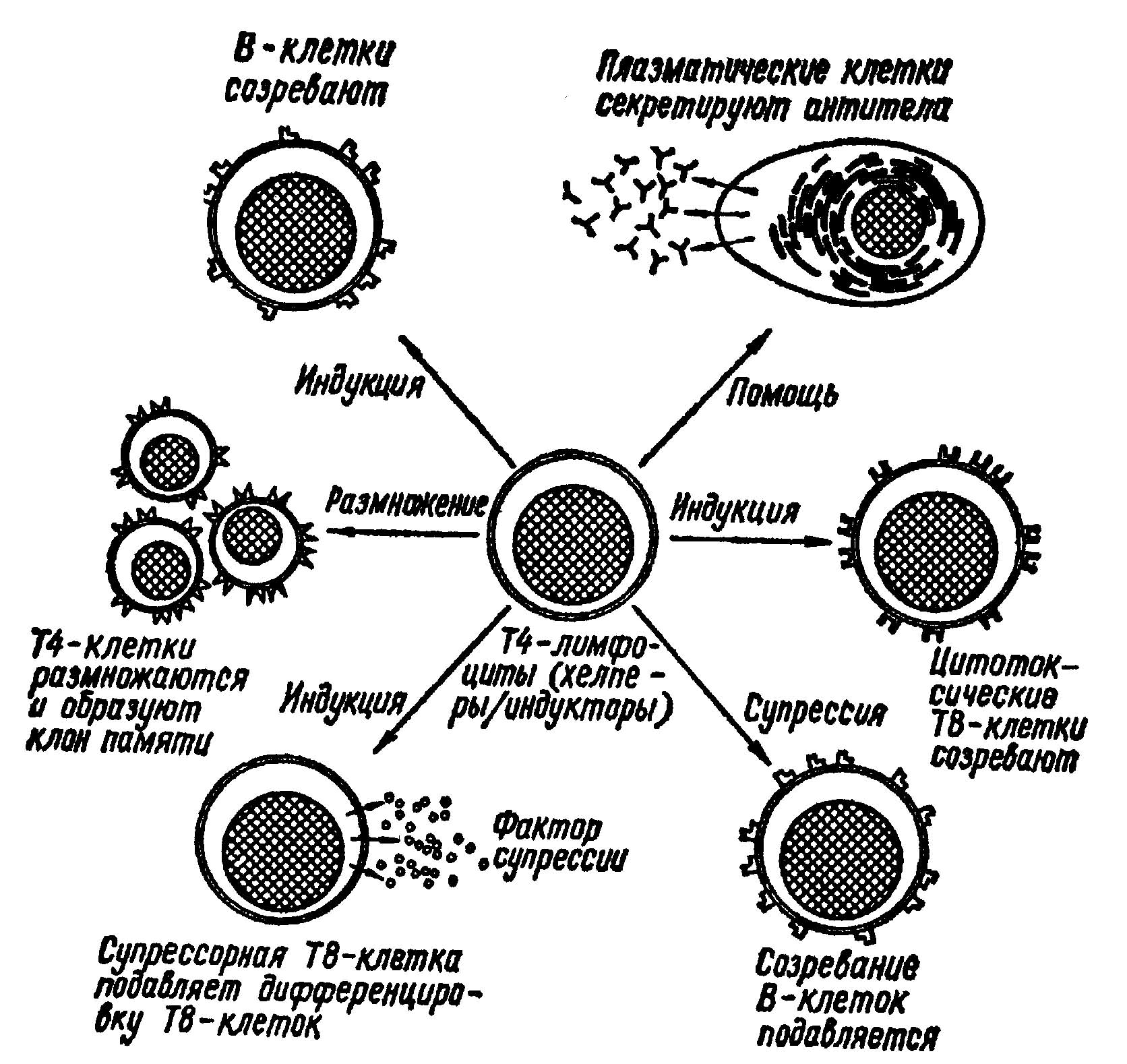
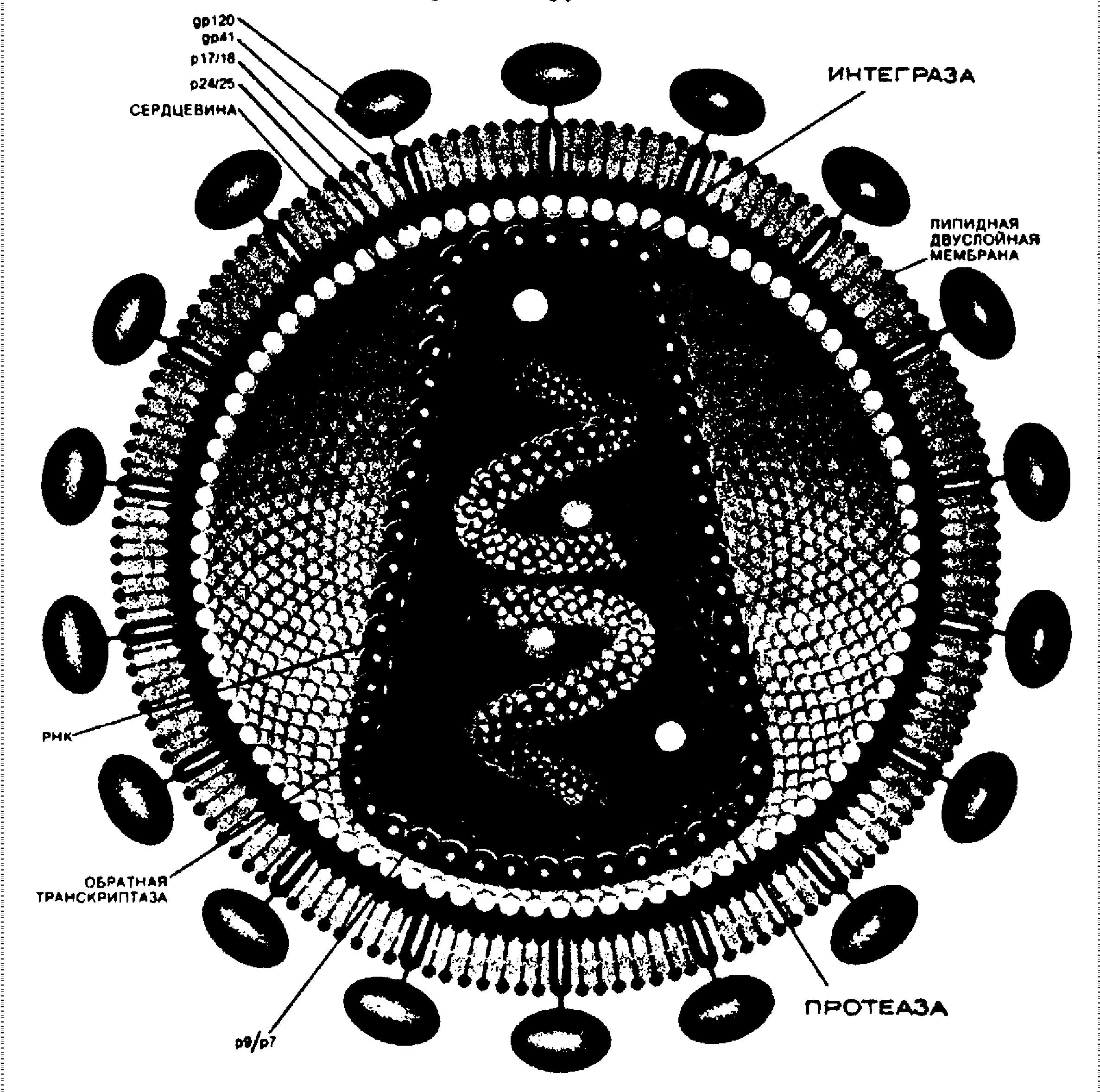
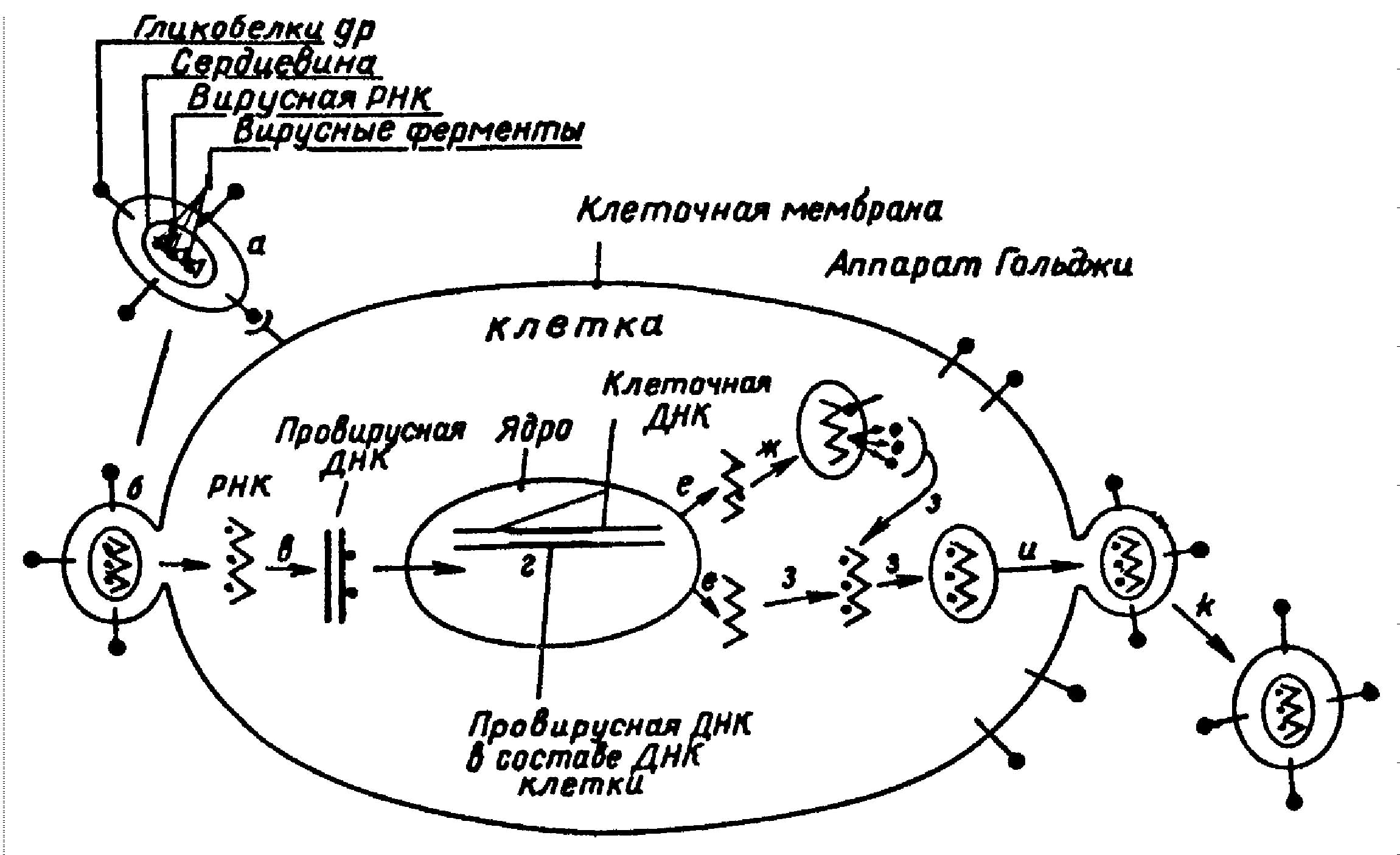
... (По-английски AIDS – acquired immunodeficiency syndrome). Далее началось период быстрого прогресса в исследованиях СПИДа, значительные новости появлялись практически каждый год. В решение проблемы, связанных со СПИДом, были вовлечены специалисты многих разделов наук: вирусологи, иммунологи, цитологи, эпидемиологи, врачи и др. Уже в 1982 – 1983 гг. была выявлена общая картина болезни, установлено ...
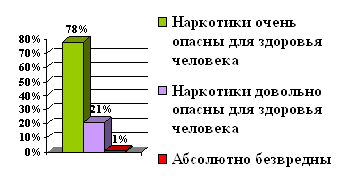
... гимназии №40 Октябрьского района г. Барнаула, проведённого в мае 2004 года) 3.1. Программа конкретного социологического исследования зависимости поведенческой культуры подростков от уровня информированности по проблеме ВИЧ/СПИД Проведенный теоретический анализ позволил сформировать основу для проведения эмпирического социологического исследования. Объектом данного исследования является ...
... 11. Обработка результатов. Поведя социологическое исследование, мы пришли к следующим выводам: 1) Гипотеза 1 о том, что большинство опрошенных считают проблему ВИЧ/СПИДа, глобальной проблемой современного общества, которая касается каждого, подтвердилась, так как 90% опрошенных на первый вопрос ответили, что проблема ВИЧ/СПИДа глобальная проблема, касающаяся каждого. 2) Гипотеза 2 о том, что ...
... регламентирующих отдельные аспекты деятельности в сфере ВИЧ/СПИДа, знание которых, наряду с национальным и международным законодательством по этой проблеме является обязательным при проведении этической экспертизы проектов, затрагивающих контингенты ВИЧ/СПИДа. Этические принципы Исследование, целью которого является работа с разнообразными биомедицинскими вопросами в области ВИЧ, сопряжено ...
0 комментариев