к нему у катушки роговой
Е.С. Боричева, В.Ф. Бондаренко, С.И. Рудая
Изучена пластичность пищевых предпочтений моллюска катушки роговой в зависимости от срока голодания в присутствии химических сигналов будущего корма в водной среде.
У брюхоногих моллюсков хеморецепция является одним из основных видов сенсорного восприятия, без которого невозможны оценка химического состава окружающей среды, размножение, хоминг, избегание врагов, а также поиск и выбор пищи [3]. Последний связан с пищевыми предпочтениями, которые определяются как врожденными реакциями на химические сигналы, так и предварительным опытом питания тем или иным кормом [3; 4]. Измерение латентного периода ориентации Achatina fulica на запах корма показало влияние мотивационных факторов (голодания) на пластичность пищевого поведения брюхоногих моллюсков. Латентный период реакции выбора корма животными в этих экспериментах уменьшался при голодании более семи дней. Показано также, что голодные Achatina fulica в течение недели после последнего приема пищи чаще выбирали запах хорошо знакомого корма по сравнению с запахом нового. Однако влияние времени голодания на скорость выработки пищевого предпочтения у водных брюхоногих моллюсков ранее не изучалось. Неизвестно также, как может влиять на эффективность выработки пищевого предпочтения к химическим сигналам будущего корма присутствие этих сигналов в окружающей среде в период голодания моллюсков. Поэтому целью нашего исследования было изучение влияния срока голодания и химических сигналов будущего корма на скорость выработки пищевого предпочтения к нему у моллюска катушки роговой.
Материал и методикаМоллюски были собраны в водоемах Калининграда в количестве 150 особей. Работа включала две серии экспериментов, каждая из которых была проведена на 75 животных. В первой серии экспериментов оценивали изменение эффективностей экстрактов старого и нового кормов у катушки роговой после различных сроков голодания. Животных разделили на пять групп по 15 особей и поместили в отдельные емкости объемом по 8 литров. Воду в аквариумах меняли каждые двое суток. Вторая серия экспериментов отличалась от первой тем, что животные голодали в присутствии химических сигналов будущего корма. Для этого пять групп моллюсков по 15 экземпляров в каждой содержали в одном аквариуме объемом 40 литров, разделенном перегородками со множеством отверстий диаметром 5 мм на пять равных секций. Воду в общем аквариуме не меняли, а лишь доливали необходимое количество свежей воды для поддержания постоянного уровня. Температура воды во всех экспериментах была +20 – 24 ºС. Исследование длилось 64 дня, и методика оценки эффективности кормов в обеих последовательных сериях наблюдений была одинаковой.
В первые 7 дней эксперимента всех моллюсков кормили кусочками свежей моркови ad libitum. Затем животных в первом аквариуме и в первой секции общего аквариума начинали кормить листьями капусты. Раз в два дня определяли эффективность экстрактов старого (морковь) и нового (капуста) кормов. В это время в остальных четырех аквариумах первой серии опытов и четырех секциях общего аквариума второй серии опытов до начала кормления капустой катушки голодали в течение 5, 10, 15 и 20 дней соответственно. По истечении этих сроков голодания оценивали эффективность экстрактов кормов. Для приготовления водных экстрактов корнеплод моркови и лист капусты натирали на мелкой терке, к чайной ложке каждого корма добавляли по 50 мл отстоянной водопроводной воды, настаивали в течение одного часа и затем фильтровали. После этого регистрировали реакции на каждый экстракт у десяти выбранных случайным образом животных. Для этого моллюска переносили из аквариума в стеклянный стакан с 20 мл профильтрованной аквариумной воды и в течение трех минут подсчитывали суммарное число скребковых движений радулы (ЧСД). Эта величина считалась фоновой ЧСД. Затем в стакан с помощью пипетки добавляли 1 мл экстракта и снова определяли ЧСД, которую считали вызванной. Затем определяли среднюю эффективность экстракта каждого корма в стимуляции ЧСД по формуле:
S ЧСДвызв. – S ЧСДфон./ n,
где n = 10.
Уровни значимости влияния водных экстрактов кормов на ЧСД моллюсков и достоверность различий между эффективностями старого и нового кормов по дням эксперимента оценивали с помощью t-критерия Стьюдента [2].
Таким образом, за 64 дня было проведено 52 измерения эффективности кормов, каждое на основании 20 наблюдений с учетом контроля.
РезультатыВ первой серии экспериментов регистрировали эффективности экстрактов старого и нового кормов в стимуляции ЧСД у катушки роговой в отсутствии химических сигналов нового корма. На рисунке 1 показано изменение эффективностей этих экстрактов в группе питавшихся животных и в группах со сроками голодания 5, 10, 15 и 20 дней.
В первом аквариуме, в котором моллюски без голодания перешли на новый корм, в первый и второй дни кормления им отдавали предпочтение старому корму – моркови. Причем в первый день экстракт нового корма даже тормозил пищевое поведение катушки роговой. Однако на четвертый день питания новым кормом разница эффективностей экстрактов старого и нового кормов уже была статистически недостоверной. Во втором аквариуме животные, несмотря на 5 дней голодания, не предпочитали химические сигналы какого-либо из кормов. В третьем аквариуме (10 дней голодания) предпочтение животными экстракта нового корма становится статистически достоверным только на шестой день питания этим кормом. В четвертом аквариуме (15 дней голодания) животные начали предпочитать экстракт нового корма уже на четвертый день питания. В пятом аквариуме моллюски, голодавшие в течение 20 дней, предпочитали экстракт нового корма при первом же предъявлении.
Во второй серии экспериментов наблюдали за изменением эффективностей старого и нового кормов в зависимости от сроков голодания в присутствии химических сигналов будущего корма (рис. 2).
В первой секции общего аквариума моллюски не голодали, а сразу после прекращения питания морковью были переведены на новый корм – капусту. На второй и четвертый дни кормления новым кормом разница в эффективностях экстрактов кормов была статистически недостоверной.
В секциях аквариума с 5 и 10 днями голодания моллюски не отдавали предпочтения экстракту нового корма. Только после 15 дней голодания моллюски стали статистически достоверно его предпочитать (рис. 2). Это предпочтение экстракта нового корма наблюдалось и в группе животных, голодавших в течение 20 дней.
Обсуждение результатовРезультаты первой серии экспериментов показывают, что увеличение срока голодания сокращает время выработки у животных ассоциаций химических сигналов, сопровождающих питание, с потребляемым кормом. Голодание в течение двадцати дней изменяет функциональное состояние животных таким образом, что они после первых же суток питания новым кормом начинают отличать его экстракт от экстракта прежнего корма. Jager [5] в своей работе по изучению влияния голодания на ЧСД радулы пресноводного легочного моллюска прудовика обыкновенного отмечает, что голодание в течение 3 – 6 дней повышает ответ на раствор сахарозы у этого животного по сравнению с голоданием в течение 9 дней или отсутствием голодания. В нашем эксперименте при максимальном сроке голодания в 20 дней средняя пищевая реакция на экстракт нового корма выше таковой при голодании в 5, 10, 15 дней или при отсутствии голодания. По-видимому, это различие объясняется тем, что катушка роговая легче переносит голодание, чем прудовик обыкновенный, и угнетение пищевой активности, вызванное голоданием у этого вида моллюсков, наблюдается в более отдаленные сроки.
Во второй серии экспериментов наблюдали такую же закономерность во влиянии голодания на выработку пищевого предпочтения к новому корму, что и в предыдущей серии. Присутствие же химических сигналов будущего корма в окружающей моллюсков среде во время голодания не влияло положительно на эффективность выработки предпочтения к будущему корму после начала питания им. Так, в первой серии эксперимента достоверное предпочтение экстракта нового корма наблюдалось после 10 дней голодания на шестой день кормления этим кормом. Во второй серии экспериментов моллюски стали отдавать предпочтение экстракту нового корма лишь после 15 дней голодания на второй день питания новым кормом. Это позволяет предположить, что химические стимулы при отсутствии питания, даже будучи сигналами потенциального корма, не воспринимаются моллюсками как химические сигналы пищи. Можно предположить, что этот вид относится к тем брюхоногим моллюскам, для которых существует достаточно широкий спектр потенциальных пищевых объектов и чьи хемосенсорные предпочтения в каждый момент времени преимущественно определяются потребляемым кормом. Подобная пластичность в изменении хемосенсорных пищевых стимулов была продемонстрирована у виноградной улитки Helix pomatia [6], африканской улитки Achatina fulica [4] и катушки роговой Planorbarius corneus [1]. Первые два вида наземных моллюсков ориентировались при поиске пищи главным образом на запах того корма, которым в это время питались (морковь, картофель или огурец). Аналогичным образом происходит хемосенсорная адаптация и у катушки роговой в водной среде с использованием водных экстрактов растительных кормов.
Список литература1. Бондаренко В.Ф. Хемосенсорная адаптация к кормам катушки роговой Planorbarius corneus (L.) // Теоретические и прикладные аспекты биологии: Межвуз. сб. науч. тр. / Калинингр. ун-т. Калининград, 1999. С. 92 – 96.
2. Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990.
3. Croll R.P. Gastropod chemoreception // Biol. Rev. 1983. Vol. 58. P. 293 – 319.
4. Croll R.P., Chase R. Plasticity of olfactory orientation to foods in the snail Achatina fulica // J. comp. physiol. 1980. Vol. 136. P. 267 – 277.
5. Jager J.C. A quantitative study of a chemoresponse to sugars in Lymnaea stagnalis (L.) // Netherl. j. zool. 1971. Vol. 21(1). P. 1 – 59.
6. Teyke T. Food attraction condicioning in the snail, Helix pomatia // Comp. physiol. 1995. Vol. 117. № 4. P. 409 – 414.
Для подготовки данной работы были использованы материалы с сайта http://elib.albertina.ru
Похожие работы
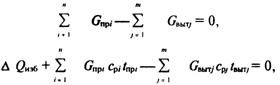
... или технологических процессов; – при выборе технического решения обеспечить малоотходность производства и максимальную эффективность использования энергоресурсов. Задачи специалиста в области безопасности жизнедеятельности сводятся к следующему; – контроль и поддержание допустимых условий (параметры микроклимата, освещение и др.) жизнедеятельности человека в техносфере; – идентификация ...
... клинического психолога. Проблемы с кожей делают людей злопамятными, а с печенью – обидчивыми. Страдающие хроническими кожными болезнями типа псориаза или дерматитов в период обострения нередко впадают в депрессии с оттенком стеснительности. Болезненно воспринимают внимание к своей внешности. Становятся очень подозрительны к людям и не выносят критики в свой адрес. При язве желудка или других ...
... ревматизма обусловила значительное снижение заболеваемости — до 0Д8 на 1000 детского населения. В разработку проблемы детского ревматизма внесли большой вклад отечественные педиатры В. И. Молчанов, А. А. Кисель, М. А, Скворцов, А. Б. Воловик, В. П. Бисярина, А. В. Долгополова и др. Эпидемиология, Установлена связь между началом заболевания и перенесенной стрептококковой инфекцией, в основном в ...
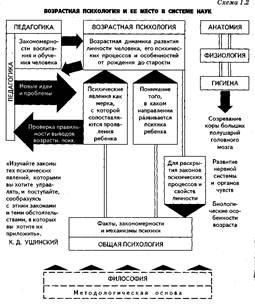

... потребовало еще больше времени. В XII в., когда статусно закреплялись другие возрасты, оно полностью отсутствовало, да и сейчас процесс его дифференциации не завершен. В частности, в англоязычной литературе по возрастной психологии такое разделение практически отсутствует до сих пор: переходный период от детства к взрослости, который связывается с хронологическими рамками от 10-12 до 23-25 лет, ...
0 комментариев