Навигация
На второй стадии образуется инициирующий комплекс
33662
знака
1
таблица
0
изображений
2. На второй стадии образуется инициирующий комплекс.
3. На третьей стадии белкового синтеза пептидная цепь удлиняется (элонгация) путем последовательного присоединения новых аминоацильных остатков.
4.На последней стадии синтез полипептидной цепи заканчивается (терминация). Сигналами терминации служат определенные участки мРНК, когда при транслокации до них доходжит очередь, синтез останавливается и готовая цепь отделяется от рибосомы.
Таким образом, синтез белка у эукариотов протекает на рибосомах, связанных с эндоплазматической сетью. Синтез белка может осуществляться также и в митохондриях, содержащих специфическую кольцевую ДНК, специфические митохондриальные формы мРНК и активирующие ферменты, а также рибосомы, сходные с рибосомами бактерий.
Вся программа химических процессов в организме записана в ДНК, молекулярном хранилище генетической информации. Расшифрована нуклеотидная последовательность ДНК и РНК, т.е. первичная структура. В 1953 году Уотсон и Крик предположили, что молекула ДНК состоит из двух правозакрученных спиральных полинуклеотидных цепей, переплетенных друг с другом и противоположно направленных. Следовательно, ДНК обладает первичной структурой – нуклеотидной последовательностью. Вторичной структурой – это 2 правозакрученные спиральные полинуклеотидные цепи, переплетенные друг с другом и противоположно направленные. Двухцепочечную структуру стабилизируют водородные связи, электростатические взаимодействия. При различных химических и физических воздействиях двухцепочечная ДНК может быть раскручена.
Данное открытие сыграло решающую роль для биологической науки 20 столетия.
Предположение о взаимодействии между азотистыми основаниями, помещенными друг напротив друга, полностью соответствует правилу Чаргаффа. Пурин всегда связан водородными связями с пиримидином (1:1). Аденин всегда связан с тимином (А=Т), а гуанин всегда связан водородными связями с цитозином (Г=Ц). Пары А=Т и Г=Ц называются комплементарными парами оснований. Более того последовательность одной из цепей полностью комплементарна последовательности второй. Противоположная полярность двух цепей в дуплексе обеспечивает пространственную ориентацию азотистых оснований.
Двухцепочечную структуру ДНК стабилизируют: водородные связи между парами оснований, электростатические взаимодействия с неорганическими противоионами, гидрофобные взаимодействия в плотно упакованных азотистых основаниях
При различных химических или физических воздействиях двухцепочечная ДНК может быть раскручена. Самый простой способ – нагревание. Повышение оптической плотности при 260 нм при нагревании известно как гиперхромный эффект.
Рассмотрим ДНК с точки зрения ее способности хранить информацию.
Четкие аргументы в пользу генетической функции ДНК были получены следующими способами. Установлено, что количество ДНК в любой клетке и организме строго постоянно и не зависит от условий внешней среды. От питания или от воздействия различных факторов, влияющих на метаболизм клетки, эта особенность ДНК вполне соответствует предполагаемым свойствам генетического материала. Содержание ДНК в клетке увеличивается с возрастанием сложности клетки и, следовательно, с возрастанием количества генетической информации в клетке.
Одним из наиболее веских аргументов в пользу генетической функции ДНК служат данные о том, что препараты ДНК, выделенные из различных тканей одного и того же организма, имеют одинаковый нуклеотидный состав. У различных видов он отличен. Таким образом, нуклеотидный состав ДНК у организма данного вида не зависит от возраста организма, условий питания и внешней среды. ДНК близких видов имеют сходный нуклеотидный состав, а эволюционно отдаленные организмы заметно отличаются один от другого по своему нуклеотидному составу.
Число пиримидиновых остатков равно числу пуриновых остатков.
Согласно модели Уотсона и Крика молекула ДНК состоит из двух правозакрученных полинуклеотидных цепей, имеющих общую ось. Две полинуклеотидные цепи в двойной спирали ДНК отличаются одна от другой как последовательностью оснований, так и нуклеотидным составом. Цепи комплементарны друг другу. Модель Уотсона и Крика объясняет, что репликация ДНК в клетке происходит в результате разделения двух цепей и последующего синтеза двух новых..
Клетки эукариотов содержат в ядре несколько или много хромосом (в зависимости от вида организма) причем в каждой хромосоме содержится одна или несколько молекул ДНК. В митохондриях эукариотических клеток содержится небольшое количество ДНК, эта ДНК отличается от ядерной. Митохондриальная ДНК – двухцепочечная, обычно кольцевая.
Денатурация двухцепочечной спирали ДНК происходит:
При экстремальных значениях рН, при нагревании, приуменьшении электрической постоянной водной среды, при обработке амидами карбоновых кислот, мочевиной. В процессе денатурации все ковалентные связи в ДНК сохраняются. Таким образом, можно сделать вывод, что ДНК представляет собой генетический материал хромосом.
Как же происходит репликация ДНК? Каковы ее ферментативные механизмы? При участии фермента ДНК полимеразы катализируется система синтеза. Эта реакция протекает только в присутствии некоторого количества преобразованной ДНК. В ее отсутствие фермент неспособен синтезировать полимер.
Преобразованная ДНК также служит и матрицей на которой фермент строит параллельную цепь ДНК, комплементарную преобразованной ДНК по составу и последовательности оснований.
Что такое репарация ДНК?
Имеются данные, что есть ферменты, способные ликвидировать разрывы, возникающие в ДНК под влиянием случайных воздействий. Такие разрывы, вероятно, реплицируются ферментом ДНК-лигазой. Также имеются данные, что репликация ДНК, чуждых данной клетке, предотвращается вследствие разрушения эндогенными нуклеазами.
Цикл элонгации протекает в 3 этапа:
Вновь поступающая аминоацил тРНК связывается с аминоацильным участком полного рибосомного комплекса, расположенного возле следующего по порядку кодона мРНК.
Для этого процесса связывания требуется ГТФ и специфический цитоплазматический белок (фактор Т).
В результате взаимодействия аминогруппы аминоацил тРНК образуется пептидная связь. Освобождающаяся транспортная РНК остается связанной в пептидильном участке. Для этого требуется фермент пептидилтрансфераза, который является частью субъединицы рибосомы.
На 3 этапе цикла элонгации пептидил тРНК физически перемещается с аминоацильного участка на пептидильный, вытесняя из него “пустую” тРНК. На этой стадии необходим специфический белок – фактор G. При каждом акте транслокации пептидил тРНК с рибосомой связывается новый комплекс фактора G с ГТФ. Одновременно с транслокацией пептидил тРНК происходит и транслокация мРНК вдоль рибосомы на один колдон.
Завершение синтеза полипептидной цепи и ее отделение от рибосомы происходит путем, отдельные этапы которого еще не изучены. Сигнал об окончании синтеза полипептидной цепи подается 3 специальными терминирующими кодонами в мРНК. Отделение полипептидил тРНК от рибосомы по достижении терминирующего кодона осуществляется при помощи специфического фактора освобождения (фактора R), который связан с рибосомой и облегчает гидролиз сложноэфирной связи между полипептидом и тРНК. Затем рибосома сходит с мРНК и после диссоциации на субъединицы может включаться в новый цикл.
Регуляция синтеза белка
Основным условием существования любых живых организмов является наличие тонкой, гибкой, согласованно действующей системы регулирования, в которой все элементы тесно связаны друг с другом. В белковом синтезе не только количественный и качественный состав белков, но и время синтеза имеет прямое отношение к многим проявлениям жизни.
Клетки живых организмов обладают способностью синтезировать огромное количество разнообразных белков, однако они никогда не синтезируют все белки. Количество и разнообразие определяется долей их участия в метаболизме. Синтез белка контролируется аллостерически. Регулируется внешними и внутренними условиями.
Жакоб и Мано сформулировали гипотезу, связывающую функционирование структурных и регуляторных генов. Эта гипотеза позволяет объяснить как индукцию, так и репрессию ферментов на основе одного механизма.Согласно этой гипотезе репрессия ферментов – основной процесс, тогда как индукция – снятие репресии.
Согласно теории Жакоба и Мано в биосинтезе белка участвуют 3 типа генов: структурные гены, ген-регулятор и ген-оператор.
Структурные гены определяют первичную структуру синтезируемого белка. Именно эти гены в цепи ДНК служат основной матрицей для биосинтеза мРНК, которая затем поступает в рибосому. Синтез мРНК на структурных генах молекулы ДНК контролируется определенным участком, называемым оператором. Он служит как бы пусковым механизмом для функционирования структурных генов.
Структурный ген кодирует аминокислотную последовательность данного фермента, синтез этого фермента происходит только в том случае, если регуляторный ген не репрессирует его, т.е. этот ген неактивен. Для объяснения репресcии было постулировано, что регуляторный ген кодирует аминокислотную последовательность специфического белка, названного репрессором. Молекула репрессора диффундирует от рибосом, где она синтезируется и образует специфический комплекс с участием ДНК, соответствующим структурному гену того фермента, который контролируется репрессором. Согласно гипотезе, молекула репрессора содержит центр связывания, специфичный для участка ДНК. Молекула репрессора обычно находится в свободном (активном “ состоянии, в котором она связывается с оператором, подавляя транскрипцию структурного гена. Если же в клетке присутствует индуктор, то образуется неактивный комплекс репрессор-индуктор, который неспособен связываться со специфическим участком ДНК (оператором) и на способен ингибировать транскрипцию структурного гена. В результате начинается транскрипция структурного гена и происходит синтез фермента. Согласно гипотезе взаимодействие между репрессором и индуктором обратимо. Таким образом, молекула репрессора должна иметь 2 специфических центра связывания: один для индуктора и другой для оператора. Центр связывания с оператором активен только в том случае, если центр связывания с индуктором свободен. Для объяснения репресии было постулировано, что в таких системах молекула репрессора в свободном состоянии неактивна и становится активной после связывания с репрессирующим метаболитом, получившим название корепрессора. Комплекс репрессор-корепрессор связывается с операторным локусом соответственного гена и, таким образом, подавляет синтез матричной РНК.
Итак гипотеза Жакоба и Мано постулирует существование двух классов белковых репрессоров: один из них используется в индуцибельном, а другой – в репрессибельных системах. Оба типа репрессоров содержат по 2 участка связывания, один из которых специфичен для операторного локуса
Жакоб и Мано обобщили свою модель регуляции белкового синтеза для случая координированной репрессии, при которой один и тот же репрессор может подавлять синтез группы ферментов и для случая координированной индукции, при которой несколько ферментов могут быть индуцированы одним и тем же индуктором.
Таким образом, концентрация некоторых ферментов возрастает при добавлении субстратов этих ферментов, это явление называется адаптацией или индукцией фермента. Концентрация ферментов может уменьшаться в присутствии конечных продуктов реакций, катализируемых этими ферментами, этот эффект называется репрессией ферментов. Индукция и репрессия ферментов взаимосвязаны. Индукция – это снятие репрессии.
Индуцибельные и репрессибельные системы контролируются генами трех типов:
Это структурные гены, кодирующие аминокислотную последовательность фермента
Регуляторные гены, кодирующие белок-репрессор
Ген-оператор , с которым специфически связывается репрессор.
Молекулы индуцирующего агента обладают способностью специфически связываться с белком репрессором. Образующийся при этом комплекс репрессор индуктор уже не может взаимодействовать с оператором, что приводит к активации транскрипции структурного гена. Группа функционально взаимосвязанных генов называется опероном. Гены, входящие в состав данного оперона, активируются или репрессируются одновременно. Каждый оперон имеет оператор.
Деятельность оперона находится под контролем гена-регулятора. Поскольку структурные гены и ген-регулятор находятся на разных участках цепи ДНК, связь между ними осуществляется при помощи вещества посредника, репрессора. Репрессор имеет сродство к оператору и обратимо соединяется с ним в комплексе. Образование комплекса блокирует синтез мРНК, а следовательно синтез белка. Таким образом, функция гена –регулятора – “запрещать” деятельность оперона, синтезирующего мРНК. Репрессор обладает способностью строго специфически связываться с определенными низкомолекулярными веществами, называемыми индукторами или эффекторами. Когда такой индуктор соединяется с репрессором, последний теряет способность связываться с геном оператором, который выходит из-под контроля гена-регулятора, и начинается синтез мРНК
В живых клетках должны функционировать точно запрограммированные механизмы, обеспечивающие нужные соотношения между количествами различных синтезируемых белков. Например, число ферментов, катализирующих реакции основного метаболического пути, должно превышать число молекул ферментов, ответственных за синтез коферментов, которые необходимы в следовых количествах. Возможность регуляции скорости синтеза различных ферментов позволяет клетке экономить РНК и белки.
Таковы теоретические модели и гипотезы о природе регуляторных процессов биосинтеза белка.
Генетический код
Одной из наиболее интригующих головоломок молекулярной биологии было выяснение вопроса каким образом четырехбуквенный “язык” ДНК, содержащий всего 4 разных азотистых основания (А,Г,Ц,Т) переводится на двадцатибуквенную речь белков через образование м РНК. Из чего состоит код? Код не может состоять из одного нуклеотида, поскольку только 4 аминокислоты могут кодироваться. Также код не может быть дуплетным, комбинация двух нуклеотидов из четырехбуквенного “алфавита” не может охватить всех АК, т.к. подобных комбинаций 16 (42=16), а в состав белка входят 20 АК.
Для кодирования всех АК белковой молекулы нужен триплетный код, где число возможных комбинаций – 64 (43=64). Генетический код для АК является вырожденным, это означает, что большинство АК кодируется несколькими кодонами. Вырожденность кода неодинакова для разных АК. Вырожденность чаще всего касается только третьего нуклеотида, в то время как почти для всех АК первые 2 нуклеотида являются общими. Она несет биологический смысл:, т.к. в процессе мутации различные АК могут быть заменены, наиболее ценные отбираются в процессе эволюции.
Другая отличительная черта генетического кода – отсутствие “знаков препинания”, т.е. сигналов, указывающих на конец одного кодона и начало другого. Другими словами код является линейным, однонаправленным и непрерывающимся. Это обеспечивает синтез в высшей степени упорядоченной последовательности молекулы белков.
Третья существенная особенность кода состоит в том, что 3 триплета не кодируют аминокислоты, они используются в качестве окончания синтеза (терминации) полипептидной цепи.
Код универсален для всех живых организмов, от инфузорий до человека. Можно заключить, что “генетический язык” в основном, одинаков для всех видов, но возможны небольшие видовые отклонения, возникшие, вероятно, в процессе эволюции и дифференцировки.
Таким образом, в ДНК имеются 4 разных основани, первоначально кодирующих всю информацию. Триплетность кода приводит к существованию 64 разных тринуклеотидных последовательностей (4*4*4). Природа кода триплетна – каждая аминокислота программируется в мРНК тремя основаниями, названными кодонами. Из простых математических вытекало, что каждая АК кодируется небольшим числом последовательно расположенных нуклеотидных остатков в цепи ДНК. Для кодирования одной АК требуется более одного нуклеотида, поскольку в ДНК обнаружено 4 типа оснований, а в белках 20 разных аминокислот. Далее, поскольку из 4 нуклеотидов (А,Г,Ц,Т) возможно лишь 16 различных сочетаний по 2 нуклеотида (42=16), кодовое “слово” для АК должно состоять более чем из двух букв. При помощи четырех разных оснований, взятых в комбинациях по три, возможно закодировать 64 (43) различные АК. Таким образом, триплетный код достаточен для кодирования всех 20 аминокислот, входящих в состав природных белков.
Характерные особенности генетического кода.
Генетический код вырожден, что означает, что почти каждой АК (кроме триптофана и метионина) соответствует более чем один кодон. Термин вырожденный, не означает неточный, поскольку один кодон не может кодировать более одной АК. Вырожденность генетического кода имеет важное биологическое значение. Существует неравномерность вырожденности кода. Например, код для серина и лейцина вырожден шестикратно, т.е. имеется 6 кодонов для серина и 6 - для лейцина, тогда как для многих других аминокислот (глутаминовая кислота, тирозин, гистидин) код характеризуется лишь двукратной вырожденностью.
Во многих случаях вырожденность затрагивает только третье основание в кодоне. Например, аланин кодируется : 2 первых основания одинаковы для всех четырех кодонов аланина. Иногда, если 2 аминокислоты имеют кодоны, в которых 2 первых основания одинаковы, то третье может быть только пурин или пиримидин.
Другая существенная особенность генетического кода состоит в том, что в нем полностью отсутствуют “сигналы” указывающие на конец и начало другого (т.е. знаки препинания). Поэтому считывание должно начинаться с определенного места молекулы матричной РНК и продолжаться последовательно от одного триплета к другому, иначе нуклеотидная последовательность во всех кодонах окажется измененной, что в свою очередь приведет к образованию белка с искаженной АК последовательностью.
Третья существенная особенность кода состоит в том, что из 64 триплетов –3 не кодируют АК. Они используются в качестве сигнала окончания синтеза (терминации полипептидной цепи).
Универсальность кода. Триплеты, кодирующие одну и туже АК одинаковы у человека, растений табака и инфузорий. Ряд экспериментов подтверждает вывод об универсальности кода. Можно заключить, что “генетический язык” в основном одинаков для всех видов, но возможны небольшие видовые отклонения, возникшие, вероятно в процессе эволюции и дифференцировки.
Вырожденность генетического кода обеспечивает организмам селективное преимущество. Если бы 20 АК кодировалось только 20 из 64 возможных кодонов, то большинство мутаций в кодирующих триплетах приводило бы к возникновению бессмысленных триплетов на кодирующих АК. При существующем генетическом коле мутация вызывает появление кодона с измененным смыслом, кодирующего другую АК или кодона синонима. Мутантный белок при этом оказывается не только функционально полноценным, но и даже полезным. Вырожденность кода способствует совершенствованию генома и его продуктов, поскольку в результате мутации происходят различные АК замены из которых в процессе эволюции отбираются наиболее ценные для выживания.
Несмотря на универсальность кода, относительное содержание четырех оснований в ДНК может существенно варьировать от вида к виду.
Как мы уже отмечали, в генетическом коде отсутствуют “запятые” между соседними кодонами, но имеются специальные знаки, отмечающие начало и конец полипептидной цепи.
Три триплета ( УАГ, УАА И УГА) вообще не кодируют АК. Они идентифицированы как сигналы терминации, т.е. окончания синтеза полипептидной цепи. В полипептидных цепях некоторых белков содержатся АК, для которых нет соответствующих “слов” в генетическом коде. Эти “некодируемые” АК являются производными “обычных” АК, которым соответствуют определенные кодоны. Эти редкие АК образуются в результате ферментативной модификации из АК, производными которых они являются уже после того, как эти аминокислоты включились в полипепдидную цепь в соответствии с кодом.
ЛИТЕРАТУРА
| 1. | Мецлер Д. Биохимия. Т. 1, 2, 3. “Мир” | 2000 |
| 2. | Ленинджер Д. Основы биохимии. Т.1, 2, 3. “Мир” | 2002 |
| 3. | Фримель Г. Иммунологические методы. М. “Медицина” | 2007 |
| 4. | Медицинская электронная аппаратура для здравоохранения. М. | 2001 |
| 5. | Резников А.Г. Методы определения гормонов. Киев “Наукова думка” | 2000 |
| 6. | Бредикис Ю.Ю. Очерки клинической электроники. М. “Медицина” | 1999 |
Похожие работы
... , вызванные динамическими му-тациями.-----------------------T-----------T-------T-----T------T------T----------------------¬ Болезнь, номер по ¦ Ген, лока-¦Триплет¦Норма¦Прему-¦Мута- ¦Литература ¦ МакКьюсику (MIM) ¦ лизация ¦ ¦ ¦тация ¦ция ¦ ¦ -----------------------+-----------+-------+-----+------+------+----------------------+ Синдром ломкой X-хро- ¦FMR1, FRAXA¦(CGG)n ...
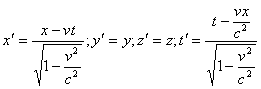
... инерциальных системах отсчета. Пространственно-временной континуум – неразрывная связь пространства и времени и их зависимость от системы отсчета. Тема 11. Основные концепции химии 1. Химия как наука, ее предмет и проблемы Важнейшим разделом современного естествознания является химия. Она играет большую роль в решении наиболее актуальных и перспективных проблем современного общества. К ...
... ", может начать по-другому регулироваться, работать в других тканях или на иных этапах развития организма. Но это – долгий путь. В какой степени он был реализован в эволюции человека, толком пока не известно. Сейчас генетики занимаются в основном подготовительной работой: проводят широкомасштабные геномные "сканирования" и составляют длинные списки "генов-кандидатов". На сегодняшний день одним из ...
... полей космоса и окружающего нас мира. Изучение информационных полей живого вещества и сфер биологических макромолекул-кристаллоидов может дать дополнительные сведения о природе и принципах организации живой формы материи. 4. Единство вещества, энергии и информации – основной принцип существования живой формы материи. Вещество, энергия и информация являются важнейшими сущностями нашего мира и ...
0 комментариев