Навигация
4. Гаметогенез.
Первинні статеві клітини — гаметобласти виникають у зародка на ранній стадії розвитку. В цей період вони ще не мають статевих особливостей і являють собою великі клітини. Вони відрізняються від інших клітин зародка вмістом глікогену та активними ферментами, серед яких важливе місце належить лужній фосфатазі. За цією ознакою їх виявляють, ставлячи відповідні гістохімічні реакції. Сперматогенез відбувається в звивистих сім'яних канальцях, у яких містяться сперматогенні клітини та клітини Сертолі, останні утворюють мікросередовище для сперматогенних клітин.
В сперматогенезі розрізняють чотири періоди: розмноження, росту, дозрівання, формування.
На стадії розмноження клітини називаються сперматогоніями. Вони межують з базальною мембраною, тобто займають периферійну частину звивистого сім'яного канальця. Сперматогонії невеликого розміру містять диплоїдний набір хромосом. Серед них існує два різновиди: одні з них мають овальне світле ядро з розсіяним у ньому хроматином. Діляться вони мітозом, збільшуючись кількісно. Це стовбурові клітини. Сперматогонії другого різновиду мають кулясте ядро, що містить зернистий хроматин. Такі сперматогонії перестають ділитися і переходять в стадію росту, перетворюючись у сперматоцити першого порядку. Вони збільшені в розмірі, зміщені в напрямі просвіту звивистого сім'яного канальця, зберігають диплоїдний набір хромосом, проходять профазу мейозу, а перед початком профази у сперматоциті першого порядку подвоюється ДНК, а також відбуваються складні зміни в перерозподілі спадкового матеріалу в ядрі, в зв'язку з цим спостерігаються такі фази: лептотени, зиготени, пахітени, дишютени. У фазі лептотени помітна ядерна мембрана, ядерце та хромосоми, що мають вигляд тонких ниток, зиготени — гомологічні хромосоми розміщуються парами, кон'югуючись за довжиною і утворюють біваленти, або діади.
У зиготенній фазі гомологічні хромосоми обмінюються генами, що забезпечує мінливість спадкового матеріалу в ряді поколінь. На стадії пахітени обидві аутосоми спіралізуються, потовщуються, стають коротшими, залишаються у тісному контакті по всій довжині. На цій стадії закінчується кон'югація хромосом. В диплотенній в кожній із кон'югуючих гомологів з'являються по парі сестринських хроматид. Хроматиди пов'язані центромерою, що перетворює біваленти в тетради. В кожній парі виникає повздовжня щілина, і у двох кон'югованих хромосомах утворюється чотири хроматиди. В тетрадах з'являються перехрести хромосом (хіазми), що свідчить про обмін гомологічними частинами між несестринськими хроматидами в тетраді. В стадії диплотени зникають сили гомологічного притягання і виникають сили відштовхування між сестринськими хроматидами. На стадії діакінезу хромосоми ще більше спіралізуються, потовщуються і відокремлюються одна від другої. За періодом росту сперматоцити першого порядку вступають в період дозрівання, який відбувається у два етапи. Після першого — редукційного поділу сперматоцита першого порядку утворюється два сперматоцити другого порядку, кожний з них одержує половину від кількості хромосом сперматоцита першого порядку. Сперматоцити другого порядку містять гаплоїдний набір хромосом. Кожна із хромосом має по дві хроматиди. Сперматоцит другого порядку менших розмірів. Перед другим поділом дозрівання у сперматоциті другого порядку не синтезується ДНК і не подвоюється хромосомний матеріал. У результаті другого — екваційного поділу дозрівання утворюється сперматида — клітина з гаплоїдним набором хромосом, кожна з яких складається з однієї хроматиди. При цьому пари хроматид, які з'єднані між собою центромерою, роз'єднуються і розподіляються між сперматидами, стають хромосомами або гонадами, сперматиди малого розміру, бідні на хроматин, містять мітохондрії, центросому, комплекс Гольджі, інші органели, а включення майже відсутні. Залежно від стадії розвитку сперматиди можуть мати округлу чи витягнуту форму. Сперматиди округлої форми розміщуються біля просвіту звивистого сім'яного канальця, а витягнутої форми — занурюються в цитоплазму клітин Сертолі, в якій вони переходять в період формування. Таким чином, у фазі дозрівання в результаті мейозу із кожного сперматоцита першого порядку виникає чотири сперматиди з одинарною кількістю хромосом. Статеві X і У-хромосоми, що характерні для сперміїв, розходяться по різних сперматидах, у зв'язку з чим Х-хромосома несе в собі жіночі ознаки, другий різновид несе в собі У-хромосому, з нього при злитті з яйцеклітиною може розвинутися організм чоловіка. Цей процес зумовлює гетерогаметність. Спосіб поділу статевих клітин, при якому дочірні клітини одержують вдвоє менше хромосом, називають мейозом, або редукційним поділом, воно перетворює диплоїдне ядро в гаплоїдне, через комбінаторику гомологів із різних пар і через кросінговер, тобто механізм, який веде до обміну генами, різко збільшує комбінаційну спадкову мінливість організмів.
Стадія формування характеризується перетворенням сперматид в спермії. При цьому їх ядра ущільнюються, набувають овальної форми, перетворюються у головки сперміїв і займають ексцентричне положення в клітині. Одночасно вони частково занурюються в цитоплазму клітин Сертолі, що зумовлює перетворення сперматид в спермії. Вважається, що клітини Сертолі забезпечують трофіку клітин сперматогенного епітелію, фагоцитують його залишки, що виникають в процесі їх дегенерації та інш. Деталі перетворення сперматиди в спермія можна виявити за допомогою електронної мікроскопії. Так, частина цитоплазми сперматиди, що містить апарат Гольджі, концентрується на апікальному кінці головки спермія. У цій зоні виникає ущільнена гранула — акробласт. Він збільшується в розмірі і у вигляді чохлика покриває ядро, в середині акробласта утворюється акросома. Центросома входить до складу шийки, вона складається із двох центріолей, зміщується в протилежний кінець сперматиди. Проксимальна центріоля прилягає до поверхні ядра, а дистальна поділяється на дві частини. Від передньої частини дистальної центріолі помітно початок формування джгутика спермія, задня половина дистальної центріолі набуває вигляду кілечка, зміщуючись по джгутику, це кілечко визначає задню межу середньої частини спермія.
Цитоплазма в міру росту хвоста сповзає з ядра і концентрується у зв'язуючій частині. Мітохондрії розміщуються спіралевидно між проксимальною центріо-лею та кілечком. Цитоплазма сперматиди під час її перетворення в спермій значно редукується. В ділянці головки вона зберігається лише у вигляді тонкого шару, що покриває акросому, незначна її кількість сповзає в дистальному напрямку і утворює тонку оболонку хвостика, а його кінцева частина зовсім не має цитоплазми. Невикористана частина цитоплазми відокремлюється і розпадається у просвіті звивистого сім'яного канальця.
Таким чином, кожна початкова сперматогонія дає початок чотирьом сперміям. Сперматогенний епітелій занадто чутливий до впливу інтоксикації, авітамінозу, дії іонізуючого випромінювання. При недостатньому живленні сперматогенез послаблюється або зовсім зникає, а сперматогенний епітелій атрофується.
Овогенез — розвиток статевих жіночих клітин.
На відміну від сперматогенезу, овогенез у тварин різних класів значно різноманітніший і залежить від біології розмноження. Він перебігає протягом трьох періодів: розмноження, росту та дозрівання.
Період розмноження. Якщо розмноження сперматогоній починається після досягнення статевої зрілості і продовжується протягом усієї статевої активності чоловіка, то в овогенезі період розмноження відбувається лише у зародка і закінчується після народження дитини. Після народження кількість статевих клітин не збільшується.
Період росту. В період росту клітини називаються овоцитами першого порядку. З самого початку свого існування овоцит першого порядку вступає в профазу мейозу. В цей період, як і в сперматоцитах першого порядку, послідовно відбуваються такі фази: лептотена, зиготена, пахітена й диплотена. Після диплотени хромосоми розходяться, деспіралізуються, стають непомітними до кінця періоду росту. Овоцит першого порядку інтенсивно синтезує речовини за допомогою хромосом.
Нагромадження поживних речовин овоцитом першого порядку поділяють на превітелогенез (фаза невеликого росту) та вітелогенез (фаза великого росту). Превітелогенез характеризується ростом цитоплазми і переважним синтезом інформаційної РНК, яка пізніше буде матрицею для синтезу білків. Такий інтенсивний синтез у овогенезі інформаційної РНК пов'язаний з тим, що він іде на запас. Не більше третини його використовується безпосередньо в овогенезі, тоді як основна частина білкового синтезу починається після запліднення. У цей час в цитоплазмі утворюються органели, необхідні для синтезу, особливо мітохондрії, структури комплексу Гольджі. Вони скупчуються поблизу ядра, де поряд з орга-нелами є пухирці ендоплазматичної сітки та краплі жиру.
У фазі великого росту синтезуються запасні поживні речовини, що нагромаджуються у цитоплазмі і використовуються лише після запліднення.
Період дозрівання відбувається після проникнення в яйцеву клітину спермія. Дозрівання овоцита — це складні перетворення у ядрі. Як і при сперматогенезі, воно складається із двох послідовних поділів, які надають яйцеклітині гаплоїдного стану. Однак, в дозріванні яйцеклітини, на відміну від сперматогенезу, один первинний овоцит утворює лише одну зрілу яйцеклітину, яка повністю зберігає жовток, що утворився в період росту, і три неповноцінні клітини — полярні (редукційні) тільця. Вони містять лише ядро без цитоплазми, при цьому ядро первинного овоцита ділиться порівно між двома дочірніми клітинами, тобто між овоцитом другого порядку і першим редукційним тільцем. Подальший поділ приводить до того, що овоцит другого порядку також ділиться нерівномірно — виникає велика зріла яйцеклітина та друге редукційне тільце. В подальшому редукційні тільця гинуть. Під час другого поділу дозрівання яйцеклітина втрачає свою центросому.
5. Запліднення.
Запліднення — злиття статевих клітин чоловіка і жінки, результатом чого є відтворення диплоїдного набору хромосом, появи якісно нової клітини — зиготи — одноклітинного організму.
В процесі запліднення розрізняють три фази:
Перша фаза — дистантна взаємодія. При цьому важлива роль належить хімічним речовинами, що продукуються гаметами — гамонами. Гіногамони І — низькомолекулярні речовини небілкової природи, продукуються овоцитами, активізують рух сперміїв; гіногамони II (фертилізини) — видоспецифічні білки, викликають склеювання сперміїв при їх реакції з комплементарним андрогамо-ном II, що міститься в цитоплазмі спермія. Речовини, що продукують спермії називають андрогамонами (І, II). Андрогамони І є антагоністами гіногамонів І, пригнічують рух сперміїв.
Друга фаза — контактна взаємодія гамет і проникнення спермія в яйцеву клітину. Відбувається вона за допомогою акросоми і її ферментів — спермолізинів. Плазматичні мембрани в місці контакту зливаються і відбувається плазмогамія — злиття цитоплазми обох гамет. Ферменти, що виділяються із акросом (гіалуронідаза, протеаза) порушують променевий вінчик, розчиняють глікозаміноглікани, що склеюють фолікулярні клітини, останні зразу ж за яйцеклітиною переміщуються по матковій трубі завдяки миготливим війкам епітеліальних клітин слизової облонки.
Третя фаза — спермій проникає в прозору оболонку. У цей час виникає кортикальна реакція. Гранули глікозаміногліканів набрякають і перетворюються у вакуолі. Останні руйнуються, їх водянистий вміст під жовтковою оболонкою утворює перивітеліновий простір. Стінки порушених вакуолей перетворюються в мікроворсинки, вони проникають в перивітеліновий простір і охоплюють спермій. Головка і проміжна частина втягуються в яйцеву клітину. Утворення пери-вітелінового простору і потовщення вітелінової мембрани, перетворюється в оболонку запліднення.
Запліднення є сигналом закінчення дозрівання яйцеклітини, вона відокремлює від себе друге редукційне тільце (перше редукційне тільце відокремлюється ще до проникнення спермія в яйцеклітину).
Проникнення спермія стимулює процеси внутрішньоклітинного обміну, що пов'язано з активізацією ферментних систем яйцеклітини. Поряд з тим починається інтенсивне переміщення складових частин овоплазми з утворенням зон підвищеної концентрації жовткових гранул, органел, що називається овоп-лазматичною сегрегацією.
Після запліднення ядро спермія набрякає, збільшується в об'ємі, має гаплоїдний набір хромосом. Пронуклеуси переміщуються у центр зиготи. Центріолі, внесені сперміями, утворюють мітотичне веретено. Мембрани перед'ядрій розпадаються і хромосоми, одержані від чоловіка та жінки, розміщуються разом в екваторіальну платівку зиготи, яка й починає ділитися (дробитися).
Дроблення. В результаті запліднення яйцевої клітини об'єднуються хромосомні набори, настає інтенсивний метаболізм, що характеризується посиленням дихання та синтетичних процесів, зокрема синтезу ДНК, РНК, білків, підвищується активність рибосом, зростає фосфорний, калієвий та кальцієвий обмін, змінюється активність протеолітичних ферментів, підвищується проникність мембран, спостерігаються інші зміни. Всі ці метаболічні зрушення стимулюють мітотичне ділення зиготи, за яким настає поділ ядер і цитоплазми. Клітини, що поділилися, не ростуть, розмір їх зберігається. Кількість ДНК в ядрі подвоюється, після чергового поділу клітини зберігають диплоїдність.
Література:
1. Гистология, цитология и эмбриология / Под ред. О.В.Волкова, Ю.К.Елецкого. - М: Медицина, 1996.
2. Луцик О.Д., Іванова А.Й., Кабак К.С., Чайковський Ю.Б. Гістологія людини. - К.: Книга плюс, 2003. - 3-тє видання. - 592 с.
3. Новак В.П., Пилипенко М.Ю., Бичков Ю.П. Цитологія, гістологія, ембріологія.: Підручник. – К.: ВІРА-Р, 2001. – 288 с.
Похожие работы
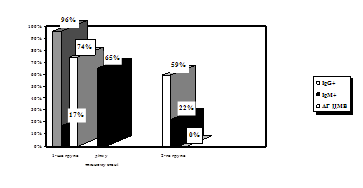
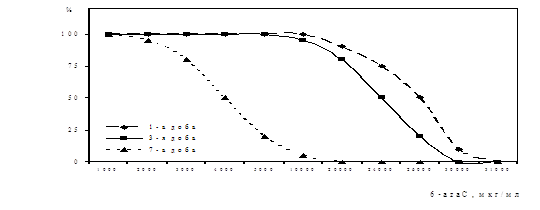
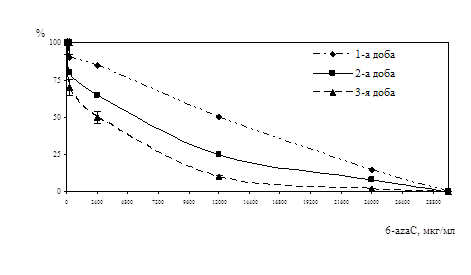
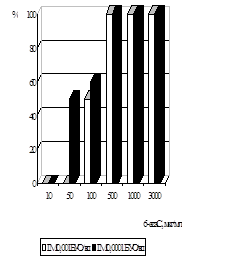
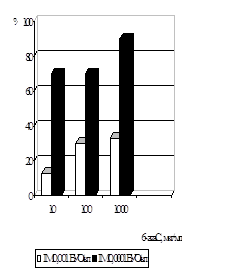
... . / Тези XIV з’їзду мікробіологів, епідеміологів та паразитологів. Полтава. - 2005, - С. 155. АНОТАЦIЯ Абдуллаєва М.В. Дослідження антивірусної активності 6-азацитидину та амізону у модельній системі: цитомегаловірус – культура клітин фібробластів легень ембріона людини. - Рукопис. Дисертація на здобуття наукового ступеня кандидата біологічних наук за спеціальністю 03.00.06 — вірусологія. — ...
... іями Провачека (реакція Вейля – Фелікса), РЗК, РНГА, РІФ. Лікування. Призначають антибіотики (Тетрациклін, Левоміцетин). Сульфаніламіди препарати протипоказані, оскільки вони посилюють ріст рикетсій. Специфічна профілактика. Проводять вакцинацію хімічною висипнотифозною вакциною. Неспецефічна профілактика – боротьба з педикульозом, рання діагностика, нагляд за контактними та іншими. 2. Ві ...
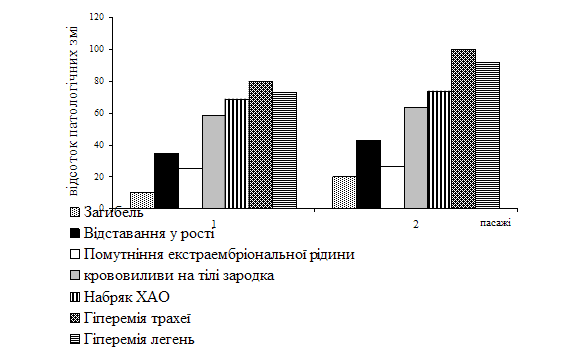


... %. На рисунку 2 представлені зародки перепелів після 2-го пасажу вакцинного штаму 1062 МПВ. Рис. 2. Зовнішній вигляд перепелиних ембріонів: а) скручування та відставання у рості ПЕ, інфікованих штамом 1062 МПВ ; б) контрольні ПЕ. Репродукцію вакцинного штаму 1062 МПВ підтверджено також за допомогою реакції імунної дифузії. Постановка РДП із гіперімунною специфічною сироваткою до ...
... до цього, однак різниця в антигенах іноді становиться причиною серйозних захворювань, які несуть загрозу нормальному розвитку вагітності і життю дитини. Частіше всього зустрічається серологічний конфлікт по резус-фактору, який може привести до небезпечних порушень, особливо до гемолітичної хвороби плоду і новонародженого. Кожна людина з моменту свого зачаття має генетично визначену групу крові. І ...
0 комментариев