Навигация
Цитратсинтазная реакция и регуляция ее скорости в мозге
82929
знаков
10
таблиц
6
изображений
5.2 Цитратсинтазная реакция и регуляция ее скорости в мозге
Скорость необратимой в физиологических условиях реакции биосинтеза лимонной кислоты находится под контролем нескольких одновременно действующих факторов. В опытах с очищенными ферментативными препаратами найдено, что АТФ является отрицательным аллостерическим модулятором цитратсин-тазы. Эффект нуклеотида обусловлен повышением константы Миха-элиса фермента для ацетилКоА.
Субстраты реакции – ацетилКоА и оксалоацетат также участвуют в регуляции активности цитратсинтазы. На основании сопоставления значений констант Михаэлиса, установленных в опытах на очищенных препаратах ферментов и реально существующих в тканях животных концентраций этих метаболитов, Кребс пришел к выводу, что in vivo основным из этих двух регуляторных факторов является концентрация щавелевоуксусной кислоты.
Таким образом, in vivo скорость цитратсинтазной реакции контролируется главным образом двумя факторами: концентрацией отрицательного аллостерического фактора фермента – АТФ и концентрацией щавелевоуксусной кислоты.
Цитратсинтазная реакция является не только важнейшим этапом ЦТК, но и компонентом системы образования ацетилхолина в холинергических нейронах. Вместе с ПДГ и цитратлиазой она обеспечивает поставку ацетил-КоА для биосинтеза нейротранс-миттера. О большом значении этой функции цитратсинтазы в мозге говорят наблюдения о том, что даже незначительные изменения активности фермента, еще не вызывающие сдвигов в энергетическом обмене, приводят к существенным нарушениям в синтезе ацетилхолина и блокируют холинергическую передачу.
5.3 Изоцитратдегидрогеназные реакции и их регуляция в мозге
Основным путем метаболизма лимонной кислоты является окисление ее в изоцитратдегидрогеназных реакциях после превращения ее под действием аконитазы в изолимонную кислоту. Активность аконитазы значительно превышает активность как цитратсинтазы, так и изоцитратдегидрогеназ и не лимитирует скорость взаимопревращения трикарбоновых кислот. В головном мозге взрослых животных до 98% нитрата подвергается дальнейшему окислению и лишь около 2% расщепляется в цитратлиазной реакции до ацетилКоА и щавелевоуксусной кислоты. В других тканях доля лимонной кислоты, подвергающаяся расшеплению цитратлиазой, может быть в несколько раз выше.
Окисление изолимонной кислоты осуществляется двумя типами изоцитратдегидрогеназ:
1. НАД-зависимым ферментом, который катализирует необратимую реакцию, протекающую исключительно в митохондриях;
2. НАДФ-специфичным ферментом, катализирующим обратимую реакцию как в митохондриях, так и в цитоплазме.
Роль НАД- и НАДФ-зависимых изоцитратдегидрогеназ в окислении изолимонной кислоты далеко не одинаково. В головном мозге основная часть субстрата окисляется по НАД-зависимому пути, поставляющему НАДН непосредственно в дыхательную цепь митохондрий и таким образом тесно связанному с поддержанием энергетического баланса клеток. Напротив, в печени, сердце и других тканях с помощью НАД-зависимой ИЦЦГ окисляется менее 10% изоцитрата, а основная масса субстрата используется в НАДФ-ИЦЦГ реакциях, особенно интенсивно протекающих в цитоплазме, где образующийся НАДФН может быть использован для разнообразных восстановительных биосинтезов.
Интересно, что явное преобладание НАД-зависимого пути окисления изолимонной кислоты в митохондриях характерно лишь для мозга взрослых животных. В то же время у растущих животных в период интенсивного липогенеза, связанного с процессами миелинизации, значительная часть изоцитрата окисляется в НАДФ-ИЦДГ-реакции и может служить источником НАДФН для биосинтеза специфических липидов мозга.
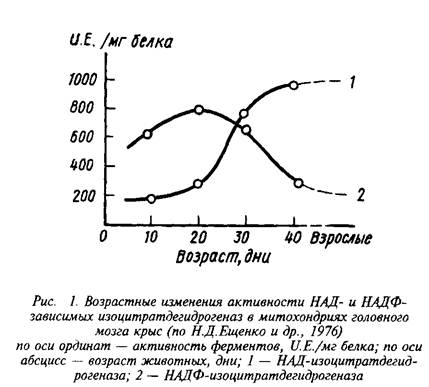
Регуляция скорости окисления изолимонной кислоты осуществляется главным образом за счет изменения активности НАД-специфичной дегидрогеназы.
В отличие от НАДФ-ИЦДГ НАД-зависимый фермент относится к числу регулируемых; положительным аллостерическим фактором служит АДФ, а АТФ, напротив, ингибирует фермент. Эффект АДФ обусловлен конформационными изменениями фермента, в результате чего возрастает в 3–10 раз его сродство к субстрату. Одновременно происходит сдвиг оптимума рН в щелочную сторону, т.е. эффектор стабилизирует НАД-ИЦДГ, обеспечивая высокую каталитическую активность в достаточно широком диапазоне рН. Установлено также, что АДФ вызывает агрегацию молекул фермента с образованием надмолекулярных форм.
Необходимо подчеркнуть, что эффективность адениннуклеотидного контроля активности НАД-ИЦДГ определяется не столько абсолютными концентрациями АТФ, АДФ и АМФ, сколько соотношением высоко- и низкоэнергетических компонентов адениннуклеотидной системы. В значительной степени эффективность адениннуклеотидного контроля НАД-ИЦДГ-реакции зависит от интрамитохондриальной концентрации ионов Са+, которые изменяют Км фермента для изоцитрата. Установлено, что низкие концентрации Са+, фумаразы, малатдегидрогеназы в мозге, как и в других тканях, превышает активность ферментов начальных стадий цикла и, следовательно, не ограничивает общую скорость цикла. Расчеты, выполненные на аналоговых вычислительных машинах, показывают, например, что на поддержание средней скорости потока метаболитов через ЦТК расходуется лишь 2–5% максимальной активности малатдегидрогеназы и около 15–20% активности сукцинатдегидрогеназы. Напротив, скорость потока метаболитов через ЦТК требует почти максимальной активности цитратсинтазы и НАД-изоцитратдегидрогеназы.
Заканчивая рассмотрение реакций ЦТК и особенностей их регуляции в головном мозге, следует кратко остановиться на значении сукцинатдегидрогеназной реакции. В отличие от других дегидрогеназ ЦТК сукцинатдегидрогеназа относится к флавин-зависимым ферментам. Она играет особую роль в энергетическом метаболизме при экстремальных состояниях, прежде всего таких, которые сопровождаются нарушениями на пиридин-нуклеотидном участке дыхательной цепи, например при облучении. При гипоксии, когда нарушается отношение между окисленными и восстановленными формами пиридиннуклеотидов, и в силу накопления НАДН возможно обращение конечных этапов ЦТК, окисление янтарной кислоты под действием сукцинатдегидрогеназы также приобретает большое значение для поддержания энергетического баланса ткани.
Из приведенных данных можно сделать вывод о следующих характерных для головного мозга особенностях в функционировании и регуляции ЦТК:
1) активность ферментов, катализирующих наиболее медленные этапы ЦТК – цитратсинтазы и НАД-изоцитратдегидроге-назы, в мозге значительно выше, чем во многих других тканях;
2) в головном мозге доминирующим механизмом регуляции скорости окисления изоцитрата является адениннуклеотидный контроль, что связано с преобладанием в митохондриях мозга НАД-зависимого пути окисления этого субстрата;
3) в головном мозге существует единый функциональный комплекс из двух ферментов – цитратсинтазы и НАД-изоцитратдегидрогеназы, обеспечивающий однонаправленное и синхронное изменение скорости наиболее медленных реакций ЦТК в зависимости от энергетических потребностей ткани, в первую очередь – от соотношения компонентов адениннуклеотидной системы;
4) на участке а-кетоглутарат–сукцинат наряду с универсальной для всех тканей последовательностью реакций в мозге возможно шунтирование с образованием в качестве промежуточного продукта биологически активной у-аминомасляной кислоты.
Похожие работы
... и постоянным для каждого индивидуума. Кровоснабжение головного мозга В норме кровоснабжение головного мозга осуществляется двумя парами - сонных и позвоночных, которые широко анастомозируют между собой концевыми ветвями, образуя на основании мозга Велизиев круг. Клинические проявления недостаточности мозгового кровообращения можно понять через сопоставление величины кровоснабжения мозга и ...
... снижение уровня неметаболического излишка СО2 и ЧСС. Итак, применение нетрадиционных природных биологически активных веществ является одним из эффективных методов дифференциро ванного повышения энергетических потенций организма спортсменов. При использовании растительных пищевых добавок в практике спорта могут быть рекомендованы следующие показания и схемы их приема: - курсовой прием продуктов ...
... нервные «приборы», посредством которых человек осуществляет анализ раздражений. Любой анализатор состоит из трех основных частей: рецептора, проводящих нервных путей и центра в коре больших полушарий головного мозга. Основной функцией рецептора является превращение энергии действующего раздражителя в нервный процесс. Вход рецептора приспособлен к приему сигналов определенной модальности (вида) – ...
... в понимание таких проблем, как соотношение духа и материи, объект-субъектный дуализм, объективность знания, редукционизм и другие. Те проблемы философско-методологическо-го характера, которые ставятся и обретают свое решение в контексте нейробиологической конструктивистской эпистемологии, Рот формулирует в виде парадоксов. Ниже приведены основные из них. ''Первый касается недостающего мира и ...
0 комментариев