Навигация
Мембранные потенциалы и их ионная природа
13949
знаков
0
таблиц
3
изображения
Мембранные потенциалы и их ионная природа
Содержание
1. Мембранные потенциалы и их ионная природа
1.1 Потенциал покоя, уравнение Нернста
1.2 Стационарный потенциал Гольдмана - Ходжкина
1.3 Уравнение электродиффузии ионов через мембрану в приближении однородного поля
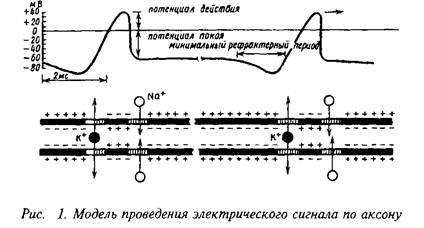
1.4 Механизм генерации и распространения потенциала действия
Список использованных источников
1. Мембранные потенциалы и их ионная природа
Мембранная теория биопотенциалов была выдвинута еще в 1902 году Бернштейном. Но только в 50-х годах эта теория была по-настоящему развита и экспериментально обоснована Ходжкиным, которому принадлежат основные идеи и теории о роли ионных градиентов в возникновении биопотенциалов и о механизме распределения ионов между клеткой и средой.
Сущность этой теории заключается в том, что потенциал покоя и потенциал действия являются по своей природе мембранными потенциалами, обусловленными полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и средой, которое поддерживается механизмами активного переноса, локализованными в самой мембране.
1.1 Потенциал покоя, уравнение Нернста
Между внутренней и наружной поверхностями клеточной мембраны всегда существует разность электрических потенциалов. Эта разность потенциалов, измеренная в состоянии физиологического покоя клетки, называется потенциалом покоя.
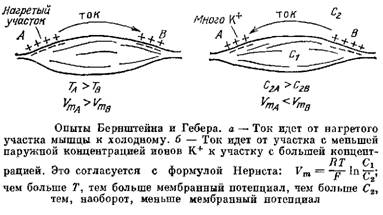
Причиной возникновения потенциалов клеток как в покое, так и при возбуждении является неравномерное распределение ионов калия и натрия между содержимым клеток и окружающей средой.
Концентрация ионов калия внутри клеток в 20 - 40 раз превышает их содержание в окружающей клетку жидкости. Напротив, концентрация натрия в межклеточной жидкости в 10 - 20 раз выше, чем внутри клеток.
Такое неравномерное распределение ионов обусловлено активным переносом ионов - работой натрий-калиевого насоса.
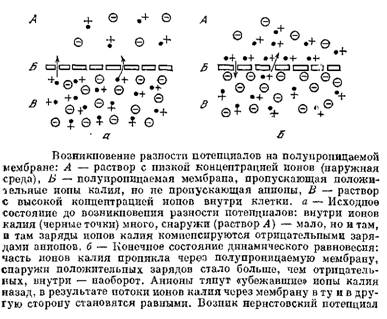
Как было установлено, возникновение потенциала покоя обусловлено, в основном, наличием концентрационного градиента ионов калия и неодинаковой проницаемостью клеточных мембран для различных ионов.
Согласно теории Ходжкина, Хаксли, Катца, клеточная мембрана в состоянии покоя проницаема, в основном, только для ионов калия.
Ионы калия диффундируют по концентрационному градиенту через клеточную мембрану в окружающую жидкость; анионы не могут проникать через мембрану и остаются на ее внутренней стороне.
Так как ионы калия имеют положительный заряд, а анионы, остающиеся на внутренней поверхности мембраны, - отрицательный, то внешняя поверхность мембраны при этом заряжается положительно, а внутренняя - отрицательно.
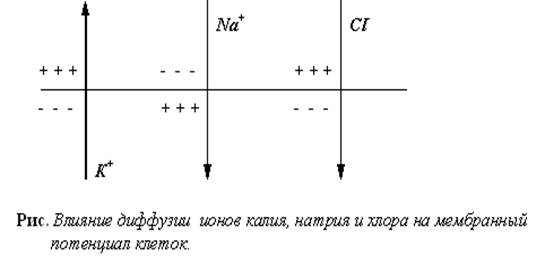
Понятно, что диффузия продолжается только до того момента, пока не установится равновесие между силами, возникающего электрического поля и силами диффузии.
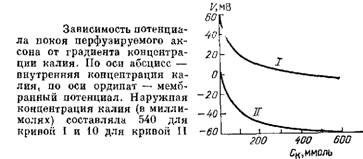
Если принять, что потенциал покоя определяется диффузией только ионов калия из цитоплазмы наружу, то его величина E может быть найдена из уравнения Нернста:
мембранный потенциал клетка электродиффузия
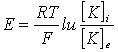 ,
,
где [K]i и [K]e - активность ионов калия внутри и снаружи клетки; F - число Фародея;T - абсолютная температура; E - изменение потенциала; R - газовая константа.
1.2 Стационарный потенциал Гольдмана - Ходжкина
Для количественного описания потенциала в условиях проницаемости мембраны для нескольких ионов Ходжкин и Катц использовали представление о том, что потенциал покоя на равновесный, а стационарный по своей природе, то есть он отражает состояние системы, когда через мембрану непрерывно идут встречные потоки ионов K+, Na+, Cl- и других.
Суммарный поток положительно заряженных частиц через мембраны равен сумме потоков одновалентных катионов минус сумма потоков одновалентных анионов.
Основной вклад в суммарный поток зарядов практически во всех клетках вносят ионы Na+, K+ и Cl-, поэтому
![]()
Наличие суммарного потока приведет к изменению потенциала на мембране; скорость этого изменения зависит от емкости мембраны. Связь между плотностью тока j ![]() , удельной емкостью С
, удельной емкостью С ![]() и потенциалом j (В) известна из курса физики:
и потенциалом j (В) известна из курса физики:
![]()
![]() ,
,
де ![]() - скорость изменения потенциала
- скорость изменения потенциала ![]() . При этом величина плотности тока " j " связана с плотностью потока одновалентных катионов Ф
. При этом величина плотности тока " j " связана с плотностью потока одновалентных катионов Ф![]() , соотношением j = Ф × F, где F - число Фарадея.
, соотношением j = Ф × F, где F - число Фарадея.
Уравнение потенциала для трех ионов имеет следующий вид:
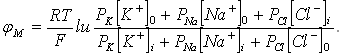
(P - проницаемость)
Это уравнение называется уравнением стационарного потенциала Гольдмана - Ходжкина - Катца.
Похожие работы
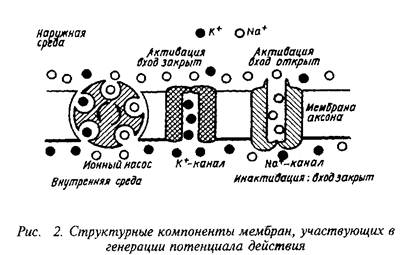
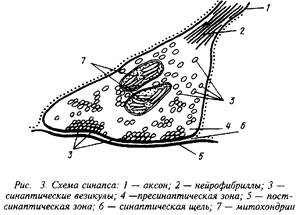
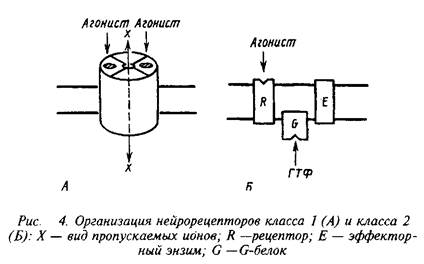
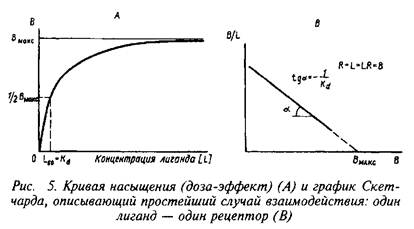
... роль нейрорецепторов сводится к созданию специфических информационных входов, организующих единый функциональный ансамбль нейронов. Именно совокупность рецепторов определяет лицо клетки и ее реакции на поступление разнообразных химических сигналов. Молекулярные механизмы, лежащие в основе модуляции эффективности синаптической передачи, в которых важную роль играют рецепторные процессы, имеют ...
... много, наружу. Вместе с ними будет выноситься наружу их положительный заряд. Внутрь через мембрану будет проходить мало ионов, так как снаружи мало калия. В результате на клеточной мембране будет возникать разность потенциалов: снаружи клетки — «плюс», а внутри — «минус». Эта разность потенциалов будет тормозить движение новых положительных заряженных ионов калия наружу, и увеличивать поток этих ...
... Формирование бислоя является особым свойством молекул липидов и реализуется даже вне клетки. Важнейшие свойства бислоя: - способность к самосборке - текучесть - ассиметричность. 1.2. Хотя основные свойства биологических мембран определяются свойствами липидного бислоя, но большинство спецефических функций обеспечивается мембранными белками. Большинство из них пронизывают бислой в виде одиночной ...
... связи. Существует и второй путь биосинтеза мевалоновой кислоты. Два автономных пути имеют значение для внутриклеточного разграничения биосинтеза холестерина необходимого для внутриклеточных нужд (синтез липопротеидов клеточных мембран) от холестерина, идущего на образование жирных кислот. В составе липопротеидов холестерин покидает печень и поступает в кровь. Содержание общего холестерина в ...
0 комментариев