Навигация
Фракционирование клеточного содержимого
67471
знак
0
таблиц
0
изображений
3.2 Фракционирование клеточного содержимого
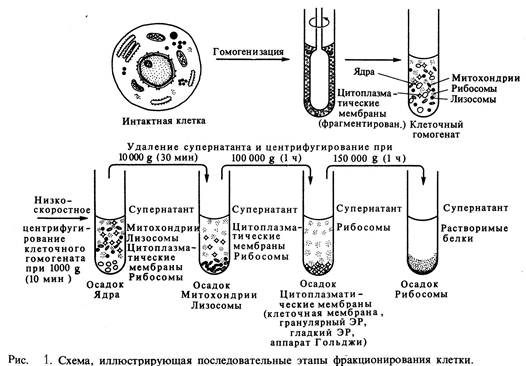
При осторожном применении методов разрушения некоторые органеллы сохраняются в интактном состоянии (ядра, минтохондрии, апарат Гольджи, лизосомы и пероксисомы). Таким образом, суспензия клеток превращается в растворимый экстракт, содержащий довольно грубую суспензию связанных с мембраною частиц, обладающих характерными размерами, зарядом и плотностью.
После того как в начале 40-х годов начали использовать препаративную центрифугу, разделение различных компонентов гомогената стало вполне реальным. Такая обработка делит клеточные компоненты по их размеру: более крупные частицы при центрифугировании движутся быстрее. Крупные компоненты экстракта, в том числе ядра или неразрушенные клетки, быстро оседают при относительно низких скоростях и образуют осадок на дне центрифужной пробирки.
Ультрацентрифуга разделяет клеточные компоненты не только по массе, но и по плавучей плотности. В этом случае образец седиментирует в круговом градиенте, образованном высококонцентрированным раствором сахарозы или хлористого цезия. Компоненты клеток опускаются по градиенту до тех пор, пока не достигнут участка, плотность раствора в котором равна собственной плотности компонентов. Дальнейшей седиментации компонентов не происходит и они "застревают" на этом уровне. Таким образом, в центрифужной пробирке возникает набор различных полос, причем полосы прилежащие к дну пробирки, содержат компоненты максимальной плавучей плотности. Данный метод настолько чувствителен, что с его помощью можно отделять немеченые макромолекулы от макромолекул, содержащих тяжелые изотопы (13С или 15N).
Фракционированные клеточные экстракты, называемые также бесклеточными системами, широко используются для изучения внутриклеточных процессов. Только работая с бесклеточными экстрактами можно установить молекулярный механизм биологических процессов, поскольку лишь в этом случае исследуемых механизм может быть изучен в чистом виде без помех, создаваемых происходящими в клетке побочными реакциями. Использование бесклеточных систем принесло первый триумфальный успех при изучении механизмов биосинтеза белка. Отправной точкой в данном случае послужил неочищенный клеточный экстракт, способный транслировать молекулы РНК в белок. После многократного фракционирования этого экстракта были получены рибосомы, РНК и различные ферменты, составляющие в совокупности аппарат биосинтеза белка. После получения отдельных компонентов в чистом виде их можно было добавлять в систему и исключать из нее и таким образом уточнять роль каждого компонента в процессе биосинтеза белка. Эта же "система трансляции in vitro" оказалась полезной для расшифровки генетического кода-с использованием в качестве матричной РНК (мРНК) искусственных полинуклеотидов известного состава.
В настоящее время различные системы трансляции in vitro применяют и для определения механизмов распределения белков по различным внутриклеточным компартментам, а также для идентификации белков, кодируемых очищенными препаратами мРНК (очистка мРНК является важным этапом в процедуре клонирования генов).
Многое из того, что мы знаем о молекулярной биологии клетки, открыто при изучении бесклеточных систем. Именно так удалось выяснить механизмы репликации ДНК, транскрипции ДНК, сплайсинга РНК, мышечного сокращения и транспорта частиц по микротрубочкам. Анализ в бесклеточных системах подразумевает полное разделение всех составляющих ее индивидуальных макромолекулярных компонентов и, в частности всех белков, входящих в систему. Методы разделения белков рассматриваются в последующих разделах.
В настоящее время хроматография является одним из методов, наиболее широко используемых для фракционирования белков. Наибольшее распространение получила распределительная хроматография.
Белки чаще всего разделяют методом хроматографии на колонках (колоночная хроматография). В этом случае смесь молекул в растворе пропускают через колонку, содержащую твердый пористый матрикс. В результате взаимодействия с матриксом различные белки проходят через колонку с различной скоростью. После того как разные белки достигнут в определенной последовательности дна колонки, их собирают отдельными фракциями. В настоящее время разработано и применяется множество матриксов различных типов, используя, которые можно делить белки согласно их заряду (ионообменная хроматография), гидрофобности (гидрофобная хроматография), размеру (хроматография гель-фильтрацией) или способности связываться различными химическими группами (аффинная хроматография).
На каждом, этапе колоночной хроматографии содержание белка в смеси увеличивается не более чем в 20 раз, и поэтому выделить из сложной смеси белков отдельный белок за один цикл практически невозможно. На долю каждого белка, как правило, приходится менее 1/1000 всего белка клетки, и для его очистки требуется последовательное использование нескольких различных типов колонок.
Гораздо более эффективен метод аффинной хроматографии (хроматография по сродству). В основе этого метода лежат биологически важные взаимодействия, происходящие на поверхности белковых молекул. Так, при ковалентном связывании субстрата фермента с матриксом, например, с полисахаридными шариками, фермент специфически удерживается матриксом и может быть элюирован (смыт) практически в чистом виде. Аффинные колонки обладают высокой степенью специфичности; за один цикл хроматографии можно добиться очень высокой степени очистки (1000-10000 раз). Применение колонок высокоэффективной жидкостной хроматографии обеспечивает высокий уровень разрешения и значительную скорость процесса. Вот почему именно этот метод чрезвычайно популярен сейчас для разделения и белков, и малых молекул.
Белки обычно несут суммарный положительный или отрицательный заряд, обусловленный наличием на их поверхности положительно или отрицательно заряженных групп аминокислот. Если белковые молекулы поместить в электрическое поле, они начинают перемещаться со скоростью, которая определяется их суммарным зарядом, а также формой и размерами. Этот феномен лежит в основе электрофореза - метода разделения смесей белков в свободных водных растворах и в твердом пористом матриксе.
В середине 60-х годов был разработан модифицированный метод электрофореза - электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН-ПААГ). При использовании данного метода белки мигрируют в инертном матриксе - полиакриламидном геле с высоким содержанием поперечных сшивок. При этом белки находятся в растворе, содержащем мощный, отрицательно заряженный детергент - додецилсульфат натрия или ДСН (SDS).
Связываясь с гидрофобными участками белковой молекулы, этот детергент вызывает развертывание белковых молекул в длинные вытянутые цепи. Развертываясь, отдельные белковые молекулы освобождаются из комплексов с белками или молекулами липидов и солюбилизируются в растворе детергента.
Каждая молекула белка связывает значительное количество негативно заряженных молекул детергента, общий заряд которых превосходит общий заряд белка. По этой причине белок после того, как будет приложено напряжение, начнет двигаться в направлении положительного электрода. Белки одного размера ведут себя сходным образом, поскольку, во-первых, их природная структура полностью нарушена ДСН так, что их форма идентична, во-вторых, они связывают одинаковое количество ДСН и приобретают одинаковый, негативный заряд. Крупные белки, обладающие большим зарядом, подвергаются действию значительных электрических сил, а также более существенному торможению. В обычных растворах эти эффекты взаимно погашаются, но в порах полиакриламидного геля, действующего как молекулярное сито, большие белки тормозятся значительно сильнее, чем малые белки. Вследствие этого сложная смесь белков делится на ряд полос, расположенных в соответствии с их молекулярной массой. Используя красители, можно выявить основные фракции полипептидов.
Метод ДСН-электрофореза белков в полиакриламидном геле значительно мощнее любого другого метода фракционирования белков, известных ранее, хотя бы потому, что может быть использован для выявления любого белка независимо от его растворимости в воде. При использовании этого метода полипептиды разделяются строго по размеру, поэтому с его помощью можно получить информацию о субъединичном составе любого комплекса и о молекулярной массе белков, образующих этот комплекс.
Метод двумерного гель-электрофореза, в котором объединены две различные процедуры разделения, позволяет идентифицировать более 1000 белков. Результаты при этом получают в виде "двумерной" белковой карты.
При работе данным методом на первом этапе белки разделяют по их заряду. Для этого образец помещают в небольшой объем раствора, содержащего неионный (незаряженный) детергент - меркаптоэтанол, и в качестве денатурирующего агента - мочевину. Диссоциированные полипептидные цепи разделяют методом изоэлектрического фокусирования, основанном на изменении заряда белковой молекулы при изменении рН окружающей среды. При изоэлектрическом фокусировании белки подвергаются электрофорезу в узкой трубочке, заполненной полиакриламидным гелем, в котором с помощью специальных буферов создается градиент рН. Под действием электрического поля каждый белок перемещается в ту зону градиента, которая соответствует его изоэлектрической точке и остается в ней. Так происходит разделение белков в одном направлении двумерного гель-электрофореза.
На втором этапе трубочка геля, содержащего разделенные белки, снова подвергается электрофорезу, на этот раз в направлении перпендикулярном тому, что на первом этапе. В этом случае электрофорез ведут в присутствии ДСН и белки разделяют по их молекулярной массе, как в одномерном ДСН-ПААГ. Неразделенными в результате остаются только те белки, которые неразличимы как по изоэлектрической точке, так и по молекулярной массе; такое сочетание встречается очень редко. Разрешение этого метода настолько велико, что позволяет разделить два практически идентичных белка, отличающихся одной заряженной аминокислотой.
В основе точной идентификации белковой молекулы лежит определение аминокислотной последовательности. Уже на первом этапе этого процесса, включающего расщепление белка на мелкие фрагменты, можно получить значительную информацию о данном белке. Так, фермент трипсин отщепляет остатки лизина и аргинина со стороны карбоксильных групп; химический реактив бромистый циан расщепляет пептидные связи, расположенные после остатков метионина. Поскольку такие специфические ферменты и реактивы расщепляют в белковой молекуле ограниченное количество связей, при их воздействии образуется смесь больших пептидов. Разделив эту смесь методом электрофореза или хроматографии, можно получить пептидную карту, характеризующую исследуемый белок. Такие пептидные карты называют иногда "фингерпринтами" (отпечатками пальцев) белка.
После разделения пептидов определяют последовательность аминокислот в каждом из выделенных пептидных фрагментов. Сперва пептид обрабатывают каким-либо реактивом, взаимодействующим только со свободной аминогруппой на его N-конце, специфически расщепляют пептидную связь. Высвобождающуюся аминокислоту идентифицируют методом хроматографии. Оставшийся пептид укорачивается на одну аминокислоту. Его также подвергают реакциям, проводимым в той же последовательности. Циклический характер этих реакций дал возможность автоматизировать весь процесс в приборах секвенаторах. На последнем этапе анализа последовательности аминокислот, полученные для пептидных фрагментов, располагают в том же порядке, как они были расположены в интактной цепи. Для этого сравнивают последовательности наборов перекрывающихся фрагментов, полученных при расщеплении одного и того же белка различными протеолитическими ферментами.
В настоящее время достаточно определить в белке 20 аминокислот, чтобы сконструировать ДНК-зонд, используемый для клонирования соответствующего гена. После выделения гена оставшаяся невыясненной часть аминокислотной последовательности белка может быть реконструирована по нуклеотидной последовательности согласно генетическому коду.
Рассмотрим два метода определения молекул внутри клеток: один из них включает использование радиоактивных изотопов, а другой - использование антител. Оба метода весьма эффективны для выявления определенных молекул в сложных смесях. Потенциально эти методы очень чувствительны и при оптимальных условиях дают возможность обнаруживать в образце молекулы, общее количество которых меньше 1000.
Радиоактивные молекулы можно использовать для исследования практически всех внутриклеточных процессов. Для этого обычно в ходе эксперимента в культуральную среду добавляют предшественник в радиоактивной форме: при этом радиоактивные молекулы смешиваются с присутствующими в клетках нерадиоактивными. Клетка использует оба типа молекул, поскольку они отличаются только массой атомного ядра. Изменение локализации радиоактивных молекул в клетке или их химические превращения можно проследить во времени. Именно этот метод дает возможность дискриминировать химически идентичные молекулы, история которых различна - например, те молекулы, которые отличаются временем синтеза. С помощью радиоактивных методов удалось определить, что почти все молекулы живой клетки постоянно разрушаются и замещаются другими молекулами.
Одна из наиболее важных областей применения радиоактивных изотопов в биологии клетки - это определение локализации радиоактивных соединений в срезах клеток или живых тканей методом радиоавтографии. При использовании этого метода живые клетки подвергаются кратковременному мечению с последующей инкубацией в течении различных промежутков времени в нерадиоактивной среде. Затем клетки фиксируют и обрабатывают для проведения световой или электронной микроскопии. Каждый приготовленный препарат покрывают тонким слоем фотоэмульсии и оставляют на несколько дней в темноте - время, в течении, которого происходит распад радиоактивного изотопа. Затем фотоэмульсию проявляют. Месторасположение радиоактивных молекул в каждой клетке можно определить по расположению темных зерен серебра.
Антителами называют белки, продуцируемые позвоночными животными для защиты от инфекции. Каждая форма антител обладает определенными участками связывания, которые предназначены для специфического узнавания молекул, стимулировавших синтез антител. Эти молекулы называют антигенами. Высокая специфичность антител в отношении антигена превращает их в мощный инструмент для исследования биологии клетки. После окрашивания антител флуоресцирующими красителями их можно использовать для определения внутриклеточной локализации специфических макромолекул с помощью флуоресцентной микроскопии. Мечение электроноплотными микрочастицами, например микросферами коллоидного золота, позволяет использовать антитела для локализации клеточных антигенов при помощи электронной микроскопии. Антитела могут выступать в роли биохимических звеньев для выявления и определения количества молекул в клеточных экстрактах и идентификации специфических белков после их разделения с помощью электрофореза в полиакриламидном геле. При связывании антител с инертным матриксом получают аффинные колонки, пригодные для выделения и очистки специфических молекул из грубых клеточных экстрактов.
Обычно антитела извлекают из сыворотки, обогащенной антителами, которую получают путем многократного введения антигена животным (например, кролику или козе). Эта антисыворотка содержит гетерогенную смесь антител, каждый тип которых был образован определенными клетками, синтезирующими антитела (В-лимфоцитами).
Проблему гетерогенности антисыворотки удалось преодолеть в 1976 г. после разработки нового метода, который произвел революцию в исследовании внутриклеточных процессов с помощью антител. Этот метод включает клонирование В-лимфоцитов, секретирующих только один определенный тип антител. Время жизни В-лимфоцитов в культуре обычно весьма ограничено. Поэтому от иммунизированных мышей получают В-лимфоциты, секретирующие отдельные виды антител, и осуществляют их слияние с "бессмертными" клетками из опухоли В-лимфоцитарного происхождения. В результате образуется гетерогенная смесь гибридных клеток, из которых отбирают гибриды, способные размножаться в культуре и синтезировать антитела определенного вида. Эти так называемые гибридомы клонируют по отдельности и получают клоны, каждый из которых является постоянным источником моноклональных антител одного типа.
Основное преимущество метода гибридом определяется возможностью получения моноклональных антител против неочищенных молекул содержащихся в сложной смеси в качестве минорного компонента. Таким образом, в принципе можно получить моноклональные антитела против любого белка, содержащегося в клетке. Каждый тип антител можно затем использовать в качестве специфического зонда как для локализации белков с помощью цитологических методов, так и для очистки белков.
Поскольку плазматическая мембрана клеток непроницаема для крупных молекул, белки, расположенные внутри живых клеток, не могут взаимодействовать с антителами, добавляемыми извне. Если такие белки необходимо связать, в цитоплазму клеток эукариот можно ввести антитела и другие молекулы, инъецируя их тонкой стеклянной пипеткой через плазматическую мембрану. Прокалываемая плазматическая мембрана имеет способность "самозапаиваться" спустя некоторое время после инъекции.
Раздел IV. Технология рекомбинантных ДНК
Этот последний раздел посвящен методам изучения структуры и функции клеточных ДНК. Классический подход подразумевает использование генетических методов, позволяющих судить о функции генов, анализируя фенотипы мутантных организмов и их потомства. Этот подход по-прежнему эффективен, но в последнее время он дополнен набором методов, которые в сумме известны как "технология рекомбинантных ДНК". Эти методы существенно расширили возможности генетических исследований, поскольку с их помощью удается проводить как прямой контроль, так и детальный химический анализ генетического материала. Используя методологию рекомбинантных ДНК, удается даже минорные клеточные белки получать в больших количествах и, следовательно, проводить тонкие биохимические исследования структуры и функции белка.
Еще не так давно, всего лишь в начале 70-х годов биохимики считали, что ДНК является наиболее сложным для исследования компонентом клетки. Чрезвычайно длинную, химически монотонную последовательность нуклеотидов в наследственном материале тогда можно было исследовать лишь с помощью косвенных методов - либо определяя структуру белка или РНК, либо с помощью генетического анализа. В настоящее время можно вырезать отдельные участки ДНК, получать их практически в неограниченном количестве и определять последовательность нуклеотидов по нескольку сот нуклеотидов в день.
С помощью этих же методов можно по желанию экспериментатора изменить выделенный ген и ввести его вновь в геном культивируемых клеток или эмбрион животного (что несколько более сложно), где этот измененный ген начинает функционировать.
Технология рекомбинантных ДНК оказала существенное воздействие на всю клеточную биологию, позволяя исследователям решать задачи, которые раньше казались неразрешимыми, например, определять функции многих вновь открытых белков и их индивидуальных доменов, расшифровывать сложные механизмы регуляции экспрессии генов у эукариот. С помощью методов генной инженерии удалось в большом количестве получить многие белки, участвующие в регуляции клеточной пролиферации и развитии. Применение этих методов должно принести успех в крупномасштабном промышленном производстве белковых гормонов и искусственных вакцин, на получение которых ранее затрачивали очень много сил и средств.
Технология рекомбинантных ДНК включает в себя набор методов - как новых, так и заимствованных из других дисциплин, например из генетики микроорганизмов. Наиболее важные среди них это:1)специфическое расщепление ДНК рестрщирующими нуклеазами, что существенно ускоряет выделение и манипуляции с различными генами; 2)быстрое секвенирование всех нуклеотидов в очищенном фрагменте ДНК, что позволяет определить точные границы гена и аминокислотную последовательность, кодируемую им;3) гибридизация нуклеиновых кислот, позволяющая выявлять специфические последовательности РНК или ДНК с большой точностью и чувствительностью на основании их способности связывать комплементарные последовательности нуклеиновых кислот; 4)клонирование ДНК: интересующий исследователя ДНК-фрагмент вводят в самореплицирующийся генетический элемент (плазмиду или вирус), который используют для трансформации бактерий. Бактериальная клетка после трансформации воспроизводит этот фрагмент во многих миллионах идентичных копий; 5) генетическая инженерия, посредством которой последовательности ДНК изменяют с целью создания модифицированных версий генов, которые затем вновь внедряют в клетки или организмы.
Похожие работы
... новые возможности, которые возникали с появлением новых методов исследования, позволяли делать открытия, радикально менявшие взгляды на патологию, начинать качественно новые этапы её развития. Патологическая анатомия использует три основных метода исследования — вскрытие трупов людей, умерших от болезней (1); микроскопические методы изучения тканей (2); эксперимент, позволяющий моделировать на ...
... недоразумения, но затем он прочно закрепился за этими маленькими желеобразными тельцами. Современное представление о клетке тесно связано с техническими достижениями и усовершенствованиями методов исследования. Помимо обычной световой микроскопии, не утратившей своей роли, в последние несколько десятилетий большое значение приобрели поляризационная, ультрафиолетовая, флюоресцентная, ...
... физических и химических характеристик мочи: анализ содержащихся в моче твердых веществ, других растворенных веществ и микроскопическое исследование осадка мочи [4]. 2.1 Правила взятия мочи у животного исследование кровь моча животное Существует три обычных метода сбора мочи, а именно, прокол мочевого пузыря, катетеризация и сбор мочи, выделяющейся из организма естественным путем. Выбор ...
... заболевания. Определение маркеров опухолей хотя и дорогой, но очень важный метод исследования, без которого в ряде случаев обойтись просто невозможно. Большинство лабораторных методов исследования требуют специального оборудования. Так, для подготовки и сохранения проб при заданной температуре, а также проведения бактериологических и серологических исследований используют термостаты, а также ...
0 комментариев