Навигация
Методы переноса генетического материала в клетки млекопитающих
25131
знак
3
таблицы
0
изображений
Методы переноса генетического материала в клетки млекопитающих
Введение
Все эксперименты по переносу генетического материала состоят из двух отдельных этапов: переноса ДНК от донора к реципиенту и отбора реципиентных клеток, которые включили генетический материал донора. Большинство соответствующих методов позволяют ввести чужеродный материал в одну клетку из тысячи, а то и десяти миллионов, поэтому для отбора нужны весьма эффективные средства. Именно эта стадия зачастую является лимитирующей в ходе эксперимента.
Первые опыты по переносу генетического материала осуществляли с помощью слияния целых клеток. Такая техника нашла применение при изучении процессов дифференцировки и канцерогенеза, однако наиболее успешно ее использовали при картировании генов человека и получении моноклональных антител. Известно, что сформировавшийся при слиянии клеток грызуна и человека межвидовой гибрид спонтанно теряет человеческие хромосомы. Как правило, утрата хромосом происходит случайным образом, и это позволяет конструировать гибридные линии клеток, в которых содержатся разные хромосомы человека. Корреляция между присутствием конкретной хромосомы человека и экспрессией генетического маркера является основой для отнесения соответствующего гена к определенной группе сцепления. Из 1300 генов человека, картированных на сегодняшний день, примерно треть локализована на конкретных хромосомах с помощью методов генетики соматических клеток. Процесс утраты хромосом у внутривидовых гибридов происходит не так быстро, как у гибридов межвидовых. При слиянии клеток мышиной миеломы с клетками селезенки формируются стабильные линии гибридных клеток. Их характеризует иммортальность, унаследованная от миеломных клеток, и способность продуцировать антитела.
Хромосомная нестабильность межвидовых гибридов, сложности при кариотипировании тетраплоидных внутривидовых гибридов ограничивали применение методов генетики соматических клеток для генетического анализа сложных фенотипов. Необходимо было научиться переносить отдельные хромосомы между соматическими клетками. Это удалось сделать с помощью мини-клеток. Несмотря на технические трудности, метод MMGT с успехом использовали для создания гибридов и последующего картирования генов, для анализа процессов дифференцировки и развития опухоли. Такие эксперименты позволили с уверенностью говорить о существовании специфических ^rans-действующих регуляторов, участвующих в формировании фенотипа дифференцированной клетки; кроме того, был подтвержден рецессивный характер гена опухоли Вильмса.
Методы слияния целых клеток и MMGT эффективны для хромосомной локализации гена, возможность более тонкого картирования этими методами ограничена, так как оно требует наличия хромосом с транслокациями и делециями. Для решения этой задачи разработаны два метода: перенос генов, опосредованный хромосомой, и перенос генов в процессе слияния облученной клетки-донора с необлученным реципиентом. Первый способ предполагает инкубацию очищенных митотических хромосом с клетками-реципиентами в присутствии фосфата кальция. При этом происходит встраивание фрагментов донорных хромосом в хромосомы клетки-реципиента. Для идентификации гибридов, содержащих нужные фрагменты ДНК донора, применяют соответствующие методы селекции. К сожалению, встроенные фрагменты хромосом зачастую претерпевают реорганизацию, кроме того, есть достоверные данные о предпочтительном проникновении в клетку и последовательностей из центромерных областей. Эти недостатки не позволяют использовать CMGT для картирования, хотя данный метод весьма эффективен для обогащения специфическими фрагментами хромосом – составной части стратегии клонирования, называемой «обратной генетикой». Госс и Харис впервые показали, что фрагменты Х-хромосомы человека, появляющиеся в его гамма-облученной клетке, можно сохранить при слиянии этой клетки с клеткой грызуна. Углубленный анализ метода IFGT показал, что полученные фрагменты реорганизованы и для них характерна та же тенденция представлять центромерные области, что и при CMGT.
Геномную ДНК можно ввести, добавляя ее очищенный препарат к реципиентным клеткам. Эта процедура также очень важна для анализа функционирования генов в клетках млекопитающих. Разработаны три способа введения ДНК: преципитация с фосфатом кальция, с помощью ретровирусного вектора и микроинъекция. Вероятно, для каждого из них существует предельный размер фрагментов ДНК, которые переносятся без повреждения. Хотя эту гипотезу не подвергали тщательной проверке, скорее всего дело обстоит именно так. Фрагменты длиной более 100 тысяч пар оснований вряд ли можно перенести без потерь, если это вообще возможно. Метод DMGT кроме функциональных исследований используют для случайного встраивания в геном селектируемых генов и для клонирования генов, при селекции которых необходима экспрессия.
1. Общие положения
1.1 Необходимые условия
Условия культивирования клеток должны обеспечивать их максимальную жизнеспособность. Это особенно важно для клеток, используемых в эксперименте по переносу, поскольку данная процедура предполагает использование потенциально токсичных веществ, таких, например, как полиэтиленгликоль.
Успех экспериментов по генетическому переносу во многом определяется эффективностью посева клеток-реципиентов. Следовательно, эффективность посева всех клеточных линий, которые будут использованы в качестве реципиентов, должна быть проверена в условиях, аналогичных экспериментальным. Линии, демонстрирующие эффективность посева менее 10%, не годятся на роль реципиента.
Все манипуляции необходимо проводить в стерильных условиях и со стерильными реагентами. Растворы следует готовить только из реактивов, предназначенных для культивирования клеток, обязательно использовать для этого бидистиллированную воду.
Похожие работы
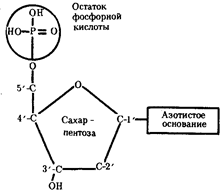
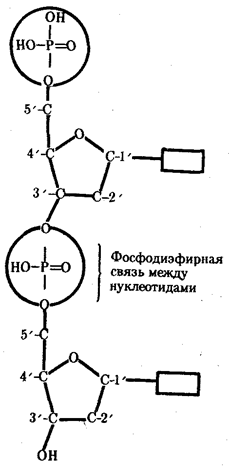
... : генный, хромосомный и геномный. На каждом из них проявляются основные свойства материала наследственности и изменчивости и определенные закономерности его передачи и функционирования. 4. Генный уровень организации генетического аппарата Элементарной функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма данного вида, ...
... , доступные генетическому анализу — это прежде всего скорость и полнота лизиса инфицированных клеток и круг бактерий-хозяев, поражаемых фагами. Широкое распространение в генетическом анализе бактериофагов получили мутанты с условным проявлением. Это мутанты, чувствительные к повышению и понижению температуры, — так называемые термочувствительные (ts) и холодочувствительные (cs). Они нормально ...
... 1972 год, когда в Стенфордском университете П. Берг и С. Коэн с сотрудниками создали первую рекомбинантную ДНК, содержавшую фрагменты ДНК вируса SV40, бактериофага и E. coli. Генетическая инженерия Одним из разделов молекулярной генетики и молекулярной биологии, который нашел наибольшее практическое приложение, является генная инженерия. Генная инженерия – это сумма методов, позволяющих ...
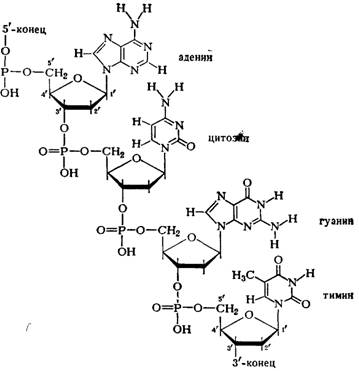
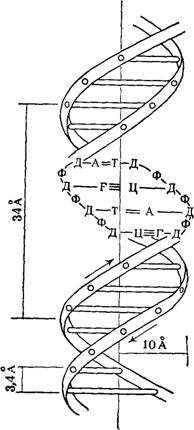
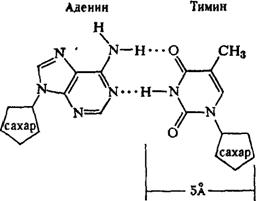
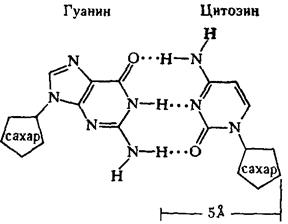
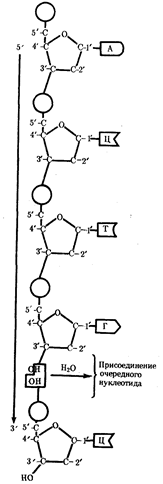
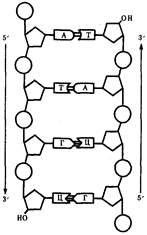
... остатки, соединяющиеся водородными связями с комплементарными основаниями матричной цепи. Рост синтезирующейся цепи происходит в направлении 3'-ОН — 3'-ОН-концам, антипараллельно матричной цепи. Репликация ДНК приводит к удвоению количества генетического материала клетки и, как правило,— к клеточному делению. Поэтому репликация происходит тем чаще, чем короче время генерации вируса или бактерии и ...
0 комментариев