Навигация
Сборка вирионов и выход их из клетки
23317
знаков
0
таблиц
0
изображений
5. Сборка вирионов и выход их из клетки
Синтез компонентов вирусных частиц в клетке разобщен и может протекать в разных структурах ядра и цитоплазмы. Вирусы, репликация которых проходит в ядрах, условно называют ядерными. В основном это ДНК-содержащие вирусы: адено-, папова-, парвовирусы, вирусы герпеса. Вирусы, реплицирующиеся в цитоплазме, называют цитоплазмэтическими. К ним относятся из ДНК-содержащих вирус оспы и большинство РНК-содержащих вирусов, за исключением ортомиксо- и ретровирусов. Однако это разделение весьма относительно, потому что в репродукции тех и других вирусов есть стадии, протекающие соответственно в цитоплазме и ядре.
Внутри ядра и цитоплазмы синтез вирусспецифических молекул также может быть разобщен. Так, например, синтез одних белков осуществляется на свободных полисомах, а других—на полисомах, связанных с мембранами. Вирусные нуклеиновые кислоты синтезируются в ассоциации с клеточными структурами вдали от полисом, которые синтезируют вирусные белки. При таком дизъюнктивном способе репродукции образование вирусной частицы возможно лишь в том случае, если вирусные нуклеиновые кислоты и белки обладают способностью при достаточной концентрации узнавать друг друга в многообразии клеточных белков и нуклеиновых кислот и самопроизвольно соединяться друг с другом, т. е. способны к самосборке.
В основе самосборки лежит специфическое белок-нуклеиновое и белок-белковое узнавание, которое может происходить в результате гидрофобных ионных и водородных связей, а также стерического соответствия. Белок-нуклеиновое узнавание ограничено небольшим участком молекулы нуклеиновой кислоты и определяется уникальными последовательностями нуклеотидов в некодирующей части вирусного генома. С этого узнавания участка генома вирусными капсидными белками начинается процесс сборки вирусной частицы. Присоединение остальных белковых молекул осуществляется за счет специфических белок-белковых взаимодействий или неспецифических белок-нуклеиновых взаимодействий.
Объединение белка с вирусными нуклеиновыми кислотами в клетке происходит спонтанно как чисто физико-химическая реакция агрегации, требующая участия дополнительных факторов (рН, ионной силы, ионов металлов, осмоса и т.п.). После того как концентрация вирусных РНК и белка достигает критического уровня; у сложно устроенных вирусов принципы самосборки обеспечивает от начала до конца морфогенез вирионов.
Для того чтобы представить этап созревания дочерних вирусных частиц у парамиксовирусов, необходимо выяснить, каким образом молекула 50S РНК, около 10 тыс. молекул белков, липидов и сахара собираются в зараженной клетке и образуют морфологически и биологически полноценную частицу. Созревание можно разделить на три этапа: 1) образование внутриклеточных нуклеокапсидов; 2) организация вирусной мембраны; 3) выход зрелой вирусной частицы из клетки с помощью так называемого почкования.
Образование нуклеокапсидов. Быстрое включение 50S РНК в капсид обусловлено сравнительно быстрым и в избытке накоплением в. зараженной клетке структурных белков нуклеокапсидов. Формирование (ассемблирование) вирионов происходит путем самосборки, которая обусловлена «узнаванием» РНК белками. Полагают, что узнающим белком является белок Р, поскольку он наиболее жестко связан с РНК в вирусных частицах. Считают, что узнаваемый участок РНК локализуется на 5'-конце молекулы. Нуклеокапсиды накапливаются в цитоплазме зараженных клеток, причем скорость образования внутриклеточных нуклеокапсидов гораздо выше, чем скорость образования вируса. Образование внутриклеточных нуклеокапсидов сопряжено во времени с биосинтезом 50S РНК.
Организация вирусной мембраны. Структурной основой вирусных и цитоплазматических мембран является двойной липидный слой. Последний у вирусной мембраны почти без изменений повторяет липидный слой плазматической мембраны клетки-хозяина. Мембраны вирусных частиц содержат наружный (гликопротеиды HN и F) и внутренний белковый слой (белок М).
В построении вирусной мембраны участвуют предсуществующие липиды клеточных мембран. Гликопротеиды вируса включаются в мембраны зараженных клеток, поэтому вскоре после их заражения в наружных мембранах клеток выявляется поверхностный вирусный антиген (гликопротеид HN, являющийся одновременно гемагглютинином, нейраминидазой и рецепторным белком).
В плазматических мембранах клеток, зараженных вирусом гриппа, обнаружены все белки липопротеидной оболочки вируса: НА (либо НА1 + НА2); NA (нейраминидаза) и М (внутренний белок вирусной мембраны). Гликопротеиды НА (гемагглютинина) и NA синтезируются в так называемых шероховатых мембранах, т.е. в мембранах, с которыми связаны синтезирующие белки рибосомы. После синтеза полипептиды транспортируются по гладким плазматическим мембранам в комплекс Гольджи, где происходит тримминг углеводных цепей. Из комплекса Гольджи гликопротеиды поступают в наружные мембраны. Происходит сборка структур, состоящих из 6 молекул гемагглютинина, вокруг одной молекулы нейраминидазы. Белок М синтезируется там же, где и гликопротеиды, но транспортируется иначе и накапливается на внутреннем листке плазмалеммы.
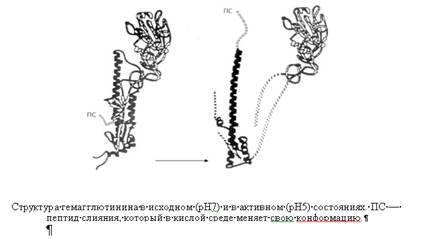
Гемагглютинин вируса гриппа синтезируется в зараженной клетке в виде предшественника, который в процессе транспорта в плазматические мембраны нарезается на две неравные субъединицы: большую и малую. Нарезание осуществляется клеточными протеазами. Степень нарезания зависит от штамма вируса и клеточной системы. При нарезании предшественника возрастает инфекционность вируса. Последняя снижена у частиц, содержащих молекулы гемагглютинина, часть которых ИЛИ все они не нарезаны. Видимо, нарезание гемагглютинина существенно для адсорбции и для стадии проникновения вируса. Обработка вирионов гриппа трипсином увеличивает эффективность адсорбции и его проникновения. Отсутствие нарезания гемагглютинина приводит к нарушению способности вирусного потомства заражать новые дочерние клетки. Нуклеокапсиды парамиксовирусов способны узнавать модифицированные М-белком участки наружных мембран зараженных клеток. Нуклеокапсиды связываются непосредственно с этими модифицированными участками. Вслед за этим начинается процесс выпячивания всего конгломерата, заканчивающийся отторжением. Этот процесс получил название «почкование». Липидосодержащие вирусы животных почкуются на плазматических мембранах клеток и содержат элементы оболочек клетки-хозяина. По сути, вирусная оболочка представляет собой модифицированную вирусными белками клеточную мембрану.
Главная роль в организации липопротеидной мембраны вируса, равно как и во включении нуклеокапсида в участок почкования, видимо, принадлежит белку М.
В связи с разнообразием структуры вирусов животных разнообразны и способы формирования вирионов, однако можно сформулировать следующие общие принципы сборки
1) у просто устроенных вирусов формируются провирионы, которые затем в результате модификаций белков превращаются в вирионы. У сложно устроенных вирусов сборка осуществляется многоступенчато — сначала формируются нуклеокапсиды или сердцевины, с которыми взаимодействуют белки наружных оболочек;
2) сборка сложно устроенных вирусов (за исключением сборки вирусов оспы и реовирусов) осуществляется на клеточных мембранах. Сборка ядерных вирусов происходит с участием ядерных мембран, сборка цитоплазматических вирусов — с участием мембран эндоплазматической сети или плазматической мембраны, куда независимо друг от друга прибывают все компоненты вирусной частицы; 3) у ряда сложно устроенных вирусов существуют специальные гидрофобные белки, выполняющие функции посредников между сформированными нуклеокапсидами и вирусными гликопротеидами. Такими белками являются матриксные белки у ряда минус-нитевых вирусов (ортомиксо-, парамиксо-, рабдовирусов);
4) сборка нуклеокапсидов, сердцевин, провирионов и вири-онов происходит не во внутриклеточной жидкости, а в специальных структурах, предсуществующих в клетке или индуцированных вирусом («фабриках»);
5) сложно устроенные вирусы для построения своих частиц используют ряд элементов клетки-хозяина, например липиды, некоторые ферменты, у ДНК-геномного SV40 — гистоны, у оболочечных РНК-геномных вирусов — актин, а в составе арена-вирусов обнаружены даже рибосомы. Клеточные молекулы несут определенные функции в вирусной частице, однако включение их в вирион может явиться и следствием случайной контаминации, как, например, включение ряда ферментов клеточных оболочек или клеточных нуклеиновых кислот.
Сборка ДНК-содержащих вирусов. В сборке ДНК-содержащих вирусов есть некоторые отличия от сборки РНК-содержащих вирусов. Как и у РНК-содержащих вирусов, сборка ДНК-содержащих вирусов является многоступенчатым процессом с образованием промежуточных форм, отличающихся от зрелых вирионов по составу полипептидов. Первый этап сборки заключается в ассоциации ДНК с внутренними белками и формировании сердцевин или нуклеокапсидов. При этом ДНК соединяется с предварительно сформированными «пустыми» капси-дами.
В результате связывания ДНК с капсидами появляется новый класс промежуточных форм, которые называются неполными формами. Помимо неполных форм с разным содержанием ДНК, существует другая промежуточная форма в морфогенезе — незрелые вирионы, отличающиеся от зрелых тем, что содержат ненарезанные предшественники полипептидов. Таким образом, морфогенез вирусов тесно связан с модификацией (процессингом) белков.
Сборка ядерных вирусов начинается в ядре, обычно — в ассоциации с ядерной мембраной. Формирующиеся в ядре Промежуточные формы вируса герпеса почкуются в перинукле-арное пространство через внутреннюю ядерную мембрану, и вирус приобретает таким путем оболочку, которая является дериватом ядерной мембраны. Дальнейшая достройка и созревание вирионов происходят в мембранах эндоплазм этической сети и в аппарате Гольджи, откуда вирус в составе цитоплазматических везикул транспортируется на клеточную поверхность.
У непочкующихся липидосодержащих вирусов (вирусов оспы) сборка вирионов происходит в уже описанных цитоплазматических «фабриках», Липидная оболочка вирусов в «фабриках» формируется из клеточных липидов путем автономной самосборки, поэтому липидный состав оболочек значительно отличается от состава липидов в клеточных мембранах
Выход вирусных частиц из клетки
Существует два способа выхода вирусного потомства из клетки: путем взрыва и путем почкования. Выход из клетки путем взрыва связан с деструкцией клетки, нарушением ее целостности, в результате чего находящиеся внутри клетки зрелые вирусные частицы оказываются в окружающей среде. Такой способ выхода из клетки присущ вирусам, не содержащим липопротеидной оболочки (пикорна-, рео-, парво-, папова-, аденовирусы). Однако некоторые из этих вирусов могут транспортироваться на клеточную поверхность до гибели клетки.
Выход из клетки путем почкования присущ вирусам, содержащим липопротеидную мембрану, которая является дериватом клеточных мембран. При этом способе клетка может длительное время сохранять жизнеспособность и продуцировать вирусное потомство, пока не произойдет полное истощение ее ресурсов.
Заключение
Разные вирусы могут вызывать самые разнообразные клеточные реакции, связанные с изменениями обмена и функции клетки. Клеточные реакции на вирусную инфекцию могут быть четырех типов. К первому из них относятся разнообразные патологические изменения, проявляющиеся угнетением синтетических процессов, нарушением функциональной активности, повреждением структуры самой клетки и ее гибелью. Такие изменения обозначаются как цитопатическая реакция на вирусную инфекцию, а способность вирусов вызывать такую реакцию — цитопатогенное действие. Второй тип клеточных реакций заключается в синтезе закодированных в клеточном геноме белков (интерферонов), обладающих антивирусной активностью. Третий тип реакции проявляется размножением вируса без видимых патологических изменений клеток (латенция). И, наконец, возможен четвертый тип, когда наличие вируса в клетке сопровождается ее пролиферацией.
Список литературы
1) Сюрнн В.Н. и др. Ветеринарная вирусология/В.Н. Сюрин, Р.В. Белоусова Н.В. Фомина —2-е над., перераб. и доп. — М.: Агропром-издат, 1991. 431 с, [8.] л, ил.: ил. — (Учебники и учеб. пособия для студентов высш. учеб. заведений).
2) Земсков М.В. и др Основы общей микробиологии, вирусологии и иммунологии. Изд. 2-е, испр. и доп. М., «Колос», 1977
3) http://collegemicrob.narod.ru/microbilogy/
4) http://vetotdel.ru/vzaimodeystvie.htm
5) http://humbio.ru/humbio/apon/0000541c.htm
Похожие работы
... , вирус Эбола и другие, способны ассоциироваться с "плотами" клеточной мембраны. Также филовирусы Эбола и Марбурга проникают в клетки путем кавеолин-опосредованного эндоцитоза, при котором белок кавеолин связан с микродоменами плазмалеммы. Механизм проникновения вирусов путем эндоцитоза, независимого от клатрина и кавеолина и опосредованного липидными площадками плазмалеммы - "плотами", главным ...
... . У штаммов HIV с мутациями, инактивирующими vif, вирионы имеют нормальный вид, содержат полный набор РНК и ферментов, но заражают клетки намного менее эффективно. 3. Этапы заражения клетки вирусом СПИДа Первый шаг любой вирусной инфекции - связывание вирусной частицы с компонентом мембраны заражаемой клетки. Для HIV роль такого рецепторного компонента играет белок, называемый антигеном CD4.( ...
... фагами, либо путем выталкивания участков цитоплазмы, либо, наконец, путем выхода отдельных вирионов или небольших их групп. Некоторые вирусы животных с трудом освобождаются из клеток в культурах in vitro; в живом организме выходу таких вирусов из клеток и их распространению способствует захват поврежденных вирусом клеток фагоцитами и их переваривание. Вирусы растений обычно не освобождаются путем ...
... ) выделяют конъюгативные и неконъюгативные плазмиды. Плазмиды могут встраиваться в хромосому бактерий- интегративные плазмиды или находиться в виде отдельной структуры- автономные плазмиды ( эписомы). Классификация и биологическая роль плазмид. Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии. Среди них- способность продуцировать экзотоксины и ферменты, ...
0 комментариев