Навигация
Роль нейромедиаторов в регуляции памяти
95237
знаков
2
таблицы
0
изображений
3. Роль нейромедиаторов в регуляции памяти
Ввиду того, что процессы памяти тесно связаны с модификацией синаптических процессов, химические передатчики нервного возбуждения должны играть здесь принципиальную роль. К настоящему времени накоплен большой экспериментальный материал, касающийся значения нейромедиаторов в процессах памяти и обучения. Полученные результаты свидетельствуют о большой значимости основных медиаторов в этих процессах, хотя конкретные формы участия каждого медиатора, по-видимому, зависят от того, какой именно тип информации запоминается.
Так, хотя известно, что способность животных к обучению, в общем, положительно коррелирует с уровнем ацетилхолина и отрицательно — с активностью холинэстеразы в мозге, тем не менее выработка одних навыков сопровождается активацией, а других — снижением активности этого фермента. В большинстве случаев, однако, ацетилхолин способствует выработке условных реакций. Показано, что снижение содержания ацетилхолина в мозге ингибиторами холинацетилазы нарушает обучение, а его повышение ускоряет выработку оборонительных навыков. В ряде исследований показано, что вещества, нарушающие обмен ацетилхолина, вызывают амнезию, а фармакологическая активация ацетилхолиновых рецепторов облегчает и ускоряет обучение и стимулирует извлечение следа из памяти.
Важная роль в регуляции процессов памяти принадлежит системе биогенных аминов мозга. В детальных исследованиях Е.А.Громовой было показано, что при выработке условных реакций с отрицательным подкреплением происходит активация норадренергической системы, а при обучении с положительным подкреплением содержание норадреналина в мозге и скорость его метаболизма снижаются. Умеренная активация норадренергических процессов стимулирует выработку реакций с болевым подкреплением, но чрезмерное увеличение содержания этого медиатора в мозге приводит к нарушению выработки всех условных реакций.
Серотонин, напротив, облегчает выработку и хранение навыков, основанных на положительном подкреплении, и отрицательно влияет на формирование оборонительных реакций. По существующим представлениям норадренергическая и серотонинергическая системы являются в значительной степени антагонистами в отношении процессов памяти, и способность к выработке тех или иных навыков зависит не столько от абсолютного уровня содержания или метаболизма того или иного медиатора, сколько от соотношения активностей этих систем. Так, нарушения, вызванные увеличением содержания серотонина, могут быть компенсированы параллельной активацией норадренергической системы и наоборот. -
По мнению Е.А. Громовой, существует реципрокность серо-тонина и норадреналина в регуляции консолидации следов памяти.
Механизм их действия заключается в свойствах этих медиаторов пролонгировать многократную циркуляцию возбуждения в нейронных системах, связанных соответственно с положительным и отрицательным восприятием информации, что является необходимым для перехода нейродинамической фазы фиксации следа в фазу структурно-химических изменений.
Сведения, касающиеся участия дофамина в регуляции процессов памяти, неоднозначны. По некоторым данным, он, как и норадреналин, может стимулировать выработку условных реакций с отрицательным подкреплением. В работах Г. Маттиеса было показано, что дофамин и его агонисты при инъекции в гиппокамп ускоряют выработку условной реакции в лабиринте с болевым подкреплением. Им же было показано, что этот медиатор стимулирует включение лейцина и фукозы в белки гиппокампальных нейронов, а также снижает фосфорилирование белка В-50 и усиливает фосфорилирование фосфоинозитолди-фосфата в клетках гиппокампа. Учитывая сказанное, есть основания считать, что дофамин участвует в регуляции синаптических процессов, связанных с фиксацией следов памяти.
Роль других медиаторов в регуляции процессов памяти изучена в меньшей степени. Из числа достаточно надежно установленных фактов можно указать на роль глутаминовой кислоты, о которой было сказано в связи с гипотезой Линча и Бодри, и на многочисленные данные, свидетельствующие о выраженном угнетающем влиянии на запоминание и обучение со стороны ГАМК.
4. Нейропептиды - регуляторы памяти
С середины 60-х годов стало известно, что некоторые олигопептиды, представляющие собой молекулы из небольшого числа аминокислотных остатков, способны модифицировать процесс обучения и влиять на степень выработки, хранения и угасания приобретенных поведенческих реакций. Среди этих пептидов оказались ранее известные гормоны, их фрагменты, а также ряд других соединений, выполняющих в организме специфические регуляторные функции. Из пептидов, относящихся к числу гормонов, наиболее выраженным действием на процессы обучения и памяти обладают гормоны гипофиза — адренокор-тикотропный и вазопрессин.
При изучении влияния АКТГ на процессы памяти было показано, что главная роль в его действии принадлежит фрагменту АКТТ4_10, который оказывает на эти процессы практически такой же эффекту как и целый гормон. Положительным влиянием на обучение обладает также и еще более короткий отрезок АХТГ4_7, хотя его действие выражено слабее, чем у АКТГ4_ J0. Более короткие фрагменты не оказывают существенного влияния на процессы обучения и памяти.
Как было выяснено, стимулирующее влияние фрагментов АКТГ на обучение не связано с собственно гормональной функцией пептида, так как фрагменты-активаторы памяти лишены такой функции. Кроме того, установлено, что эти пептиды действуют непосредственно на процессы фиксации, хранения и воспроизведения памятного следа, а не на функции, связанные с активностью и вниманием во время обучения, ибо фрагменты АКТГ оказываются эффективными при введении их либо сразу после сеанса обучения, либо перед воспроизведением приобретенного навыка.
Гормон задней доли гипофиза — вазопрессин также обладает ярко выраженным положительным влиянием на выработку условных реакций у животных и функции, связанные с памятью у людей. Будучи введенным в чрезвычайно малых дозах, он ускоряет выработку и замедляет угашение приобретенных навыков, устраняет ретроградную амнезию, улучшает воспроизведение хранящейся в памяти информации. Стимуляция вазопрессином процессов памяти также не связана с его гормональным действием, так как такое же стимулирующее действие оказывают некоторые его аналоги и фрагменты, не вызывающие свойственных вазопрессину гормональных реакций.
Есть все основания считать, что АКТГ и вазопрессин либо их фрагменты, образующиеся в организме в результате расщепления гормонов, не только стимулируют запоминание при введении их извне, но постоянно функционируют в мозге в качестве регуляторов процессов памяти. В пользу этого утверждения свидетельствует целый ряд фактов. Во-первых, введение животным иммунной антисыворотки, специфически связывающей вазопрессин либо АКТГ4-10, приводит к значительному ухудшению их обучаемости. Во-вторых, крысы, у которых в результате мутации нарушен синтез вазопрессина, способны к обучению в значительно меньшей степени, чем животные с нормальным уровнем этого нейропептида. В-третьих, доказано, что в мозге существует обширная сеть вазопрессинергических нейронов, синаптические окончания которых особенно многочисленны в тех отделах, которые принимают активное участие в процессах, связанных с памятью и обучением.
Можно, таким образом, утверждать, что вазопрессин и АКТГ и их фрагменты являются специфическими регуляторами функций центральной нервной системы, имеющих прямое отношение к процессам фиксации, хранения и воспроизведения следов памяти. Помимо этих двух соединений существует еще несколько нейропептидов, которые можно считать относительно специфичными стимуляторами запоминания и воспроизведения энграмм.
Таблица 112. Некоторые стимуляторы запоминания и воспроизведения энграммы
| Степень специфичности эффекта | Стимуляторы | |
| Общая характеристика | Вещество | |
| Относительно специфичные | Регуляторные пептиды | Вазопрессин и некоторые аналоги, дипептид pEDa, pEGa, АКТГ и некоторые аналоги. Ряд других регуляторных пептидов модулируют запоминание, но лишь в особых условиях эксперимента |
| Непептидные соединения | Пирацетам и его аналоги, некоторые ганглиозиды | |
| Агенты широкого спектра действия | Вещества, связанные с метаболизмом РНК | Оротат, метилглюкаминоротат, РНК различной полимерности и происхождения |
| Нейрости мулл торы | Фенилалкиламины, фенилалкилоиднонимины | |
| Некоторые антидепрессанты Некоторые холиномиметики и ингибиторы ацетилхолинэстеразы в ограниченном интервале доз | ||
Непептидные специфические стимуляторы памяти практически неизвестны. Единственным таким соединением, широко использующимся в клинике, эффект которого молило считать несомненным, является пирацетам. Интересно, что молекула пирацетама, химически родственная ГАМК, в то же время близка по конформации к некоторым пептидам, которые также существенно стимулируют запоминание и, в свою очередь, сходна по структуре с активными фрагментами вазоирессина.
Что касается других непептидных соединений, обладающих стимулирующим действием на процессы памяти, то их действие, скорее всего, является неспецифическим, осуществляемым благодаря общей активации деятельности ЦНС Достаточно активным стимулятором памяти является оротовая кислота, действие которой направлено на синтез уридиимоно-фосфата и, следовательно, на образование РНК.
Известно, что нейропептиды могут служить как самостоятельными медиаторами в синаптических окончаниях нейронов, так и кофакторами классических медиаторов, вместе с которыми они могут находиться либо в одной везикуле, либо в различных везикулах в пределах одной терминала.
Есть основания полагать, что такое сочетание группы активных веществ в пределах одной терминали является в большинстве случаев, если не во всех, необходимым для нормального функционирования синапса. Возможно, оно обеспечивает способность пресинаптического аппарата регулировать уровень освобождения медиатора и чувствительность к приходящим стимулам.
Известно, что нейропептиды-спутники способны значительно повышать сродство рецептора к основному медиатору. Например, вазоактивный интестинальный пептид увеличивает сродство ацетилхолиновых рецепторов к ацетилхолину более чем в десять тысяч раз. Это свойство пептидов может иметь большое значение, так как они, как правило, гораздо стабильнее, чем непептидные медиаторы. Если период полураспада у последних колеблется от нескольких долей секунды до минуты, то для многих пептидов он может составлять несколько минут и более. В результате создаются условия для того, чтобы проводимость синапса увеличивалась на время, значительно превышающее продолжительность КПМ.
Выделяющиеся в синаптических окончаниях пептиды способны влиять на описанные выше нейрохимические реакции в нейронах, которые могут быть связаны с консолидацией следов памяти. Установлено, что АКТГ, взаимодействуя с постсинаптическими рецепторами, увеличивает образование внутриклеточной цАМФ. Результатом этого может быть подавление фосфорилирования упоминавшегося синаптосомального белка В-50. Следующим звеном в цепи возникающих реакций будет являться снижение уровня фосфорилирования фосфоинозитидов, уменьшение отрицательного заряда мембраны и изменение состояния ионных каналов. Известно, что вазопрессин также оказывает влияние на уровень фосфорилирования белка В-50. Следовательно, кроме кратковременных эффектов нейропептидов-спутников, связанных с их действием на сродство рецепторов к основному медиатору, можно представить себе их более длительное действие на поляризацию других, более отдаленно расположенных мембран и на синаптическую проводимость.
Действие нейропептидов на постсинаптические рецепторы не обязательно должно быть обусловлено их локализацией в одном синаптическом окончании с непептидными медиаторами. Возможен и такой вариант, когда в районе одного постси-наптического рецептора расположены в непосредственной близости синаптические окончания двух нейронов, один из которых выделяет классический, а другой — пептидный медиатор. Оба медиатора реагируют с соседними участками постсинаптической мембраны таким образом, что пептид изменяет ее реакцию на выделение основного медиатора. В этом случае реакция постсинаптической клетки на этот медиатор в большой мере будет зависеть от того, пришел ли одновременно импульс или серия импульсов от пептидергического нейрона,
Изложенные предположения о роли нейропептидов в процессах нейронной пластичности и памяти не обязательно должны относиться исключительно к вазопрессину и АКТГ, т.е. тем пептидам, которые несомненно способны стимулировать память при их введении в организм. Вероятно, не все введенные пептиды могут, не разрушаясь, достигать тех клеток мозга, где они способны оказать свое действие. Кроме того, в том случае, когда пептид вводят системно, т.е. внутривенно, внутрибрюшинно, подкожно или внутримышечно, его действие распространяется сразу на весь организм и специфическое влияние на процессы обучения и памяти может маскироваться другими эффектами. Вот почему количество известных нейропептидов-стимуляторов памяти, скорее всего, будет постепенно увеличиваться.
Уже сейчас выявлены соединения, обладающие неизвестной ранее для них способностью стимулировать процессы запоминания и воспроизведения энграмм. Кроме того, установлено, что некоторые фрагменты и искусственно синтезированные аналоги природных пептидов часто обладают более выраженным действием, чем сами соединения, обнаруживаемые в организме. Так, выяснилось, что аналог АКТТ4_7 — MEHFKF обладает активностью, в тысячу раз превышающей активность самого АКТГ. Получен также аналог АКТГ4_7, образующийся присоединением к нему трех природных аминокислот, — MEHFPGP, который по активности не уступает исходному пептиду, но обладает значительно более продолжительным действием. Такое увеличение активности или пролонгирование эффекта объясняется тем, что присоединение дополнительных аминокислотных остатков к молекуле пептида или замена их внутри этой молекулы делает ее более устойчивой к метаболической деградации, в результате чего нейропептид достигает клеток-мишеней в больших концентрациях, чем это имеет место при введении природного соединения.
Уже упоминались случаи, когда фрагмент нейропептида оказывается активнее, чем исходное вещество. Так, было показано, что при внутримозговом введении фрагмента вазопрессина NCPGa его дозы, требуемые для стимуляции обучения, оказываются на несколько порядков ниже, чем для целого вазопрессина. Авторы этих исследований считают, что в регуляции процессов памяти участвует не вся молекула вазопрессина, а лишь фрагмент, отщепляющийся от молекулы благодаря специфике метаболизма вазопрессина в ЦНС.
Похожие работы
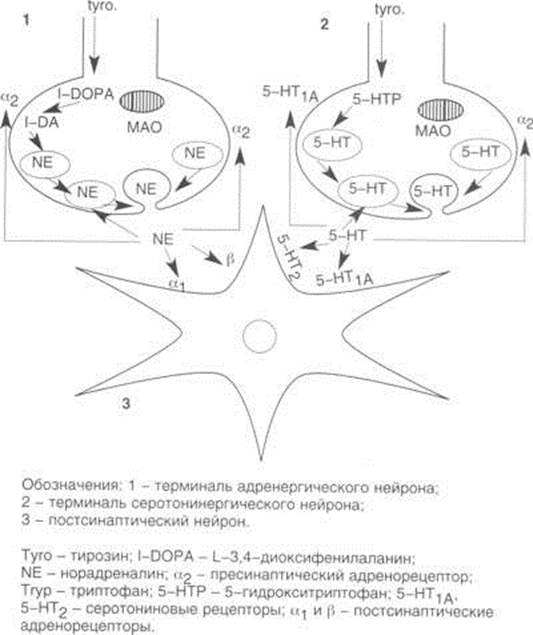
... уровне взаимодействия норадренергических и серотонинергических нервных окончаний с постсинаптическим нейроном. Общая характеристика и классификация антидепрессантов Современная классификация антидепрессантов, основанная на особенностях механизма их действия, фармакологического и нейрохимического профиля, принятая в экспериментальной и клинической психофармакологии, представлена в табл. 1. ...
... информации. Отбор информации осуществляется благодаря реакции внима- ния,рассмотренной ранее.Остановимся теперь более подробно на механизмах формирования энграммы. Важный вклад в создание современной теории памяти внес канадский физиолог Хебб.Он предположил,что внешний стимул сразу приводит к образованию лабильного следа памяти в моз- ге,который вскоре исчезает.Переход информации в долговремен- ...
... Хиден высказал предположение, что именно молекула РНК является главным нейрохимическим субстратом памяти. Опыты по изучению активности головного мозга в процессах запоминания и воспроизведения. Ключи к разгадке феномена памяти — в активности нашего головного мозга. Запоминание и узнавание уже знакомых объектов осуществляется задней и передней областями коры головного мозга. Человек обладает ...
... перекрест рук» А.Р. Лурия в общей структуре латеральных профилей мужчин и женщин в норме. Обоснованы принципы подхода к проблеме связи вариантов латеральных профилей с индивидуальными различиями с позиций дифференциальной психофизиологии, приведены нейрофизиологические и нейрохимические объяснения выявленным связям между индивидуальными профилями латеральности и некоторыми особенностями реализации ...
0 комментариев