Навигация
Проблема переноса памяти
95237
знаков
2
таблицы
0
изображений
5. Проблема переноса памяти
Предположение, что следы памяти или выработанные навыки могут быть закодированы в структуре каких-то химических соединений и переноситься с этими соединениями от одного индивида к другому, возникло на рубеже 50-60-х годов после исследований МакКоннела, проведенных на червях планариях. Безусловной реакцией планарии является сокращение тела в ответ на электроболевой раздражитель. После многократного сочетания такого воздействия со вспышкой света у червей вырабатывалась условная реакция на свет. Вслед за этим планарию разрезали на две половины и выжидали месяц, пока каждая половина не регенерирует до целой особи. После этого у всех регенерировавших таким образом планарий вновь вырабатывали ту же условную реакцию. При этом выяснилось, что для выработки реакции требуется время, в три раза меньшее, чем при первоначальном обучении, независимо от того, из какого конца — головного или хвостового — происходила регенерация. Результаты заставляли предполагать, что приобретенная условная реакция кодируется какими-то химическими веществами, которые могут храниться как в головной, так и в хвостовой части червя.
В дальнейших экспериментах была сделана попытка определить, какие именно химические соединения обусловливают перенос навыка. Оказалось, что если регенерация производится в растворе рибонуклеазы, ускорение выработки реакции происходит только у тех планарий, которые регенерируют из головного конца. Результаты этих экспериментов породили предположение, что переносчиком памяти является РНК. Однако дальнейшие исследования, выполненные на позвоночных животных, этого предположения не подтвердили.
Аналогичные эксперименты, проводимые главным образом на крысах, также показали, что передача некоторых форм приобретенных навыков с помощью химических агентов, скорее всего, возможна. Многие исследователи склоняются, однако, к представлению, что переносится не сам навык, а способность к его выработке. Большая группа экспериментов, проведенных почти одновременно в середине 60-х годов в разных лабораториях и на разных моделях, показала возможность ускорения обучения определенным навыкам у крыс-реципиентов, получавших гомогенат и экстракты мозга от обученных крыс-доноров. К переносимым относились такие реакции, как нахождение правильного пути в лабиринте, инструментальные рефлексы и привыкание к сильному звуковому раздражителю. Во всех этих экспериментах у реципиентов по сравнению с контрольными животными значительно быстрее вырабатывалась именно та реакция, которую приобрели доноры.
Дальнейшие исследования были посвящены попытке выяснить химическую природу одного из таких переносчиков памяти. Еще в 1965 г. Г. Унгар предположил, что фактором переноса является не РНК, как считалось до того времени, а белок. В дальнейшем это предположение находило все большее экспериментальное подтверждение.
Тот же Г. Унгар в последующих работах использовал методику так называемого пассивного избегания, состоящую в том, что крысу помещают в камеру, состоящую из двух отсеков — темного и светлого. В соответствии со своими биологическими склонностями животное заходит в темный отсек, где получает удар током. Если это болевое подкрепление запоминается, то при последующих помещениях в камеру животное избегает заходить в темный отсек. В опытах Г. Унгара после устойчивой выработки условной реакции пассивного избегания у крыс выделяли мозг и различные его фракции вводили реципиентам-мышам, которых впоследствии тестировали по той же методике. В результате было показано, что мыши, получавшие экстракты мозга обученных крыс, с самого начала избегали заходить в темный отсек.
Исследователями было получено большое количество одинаково обученных животных, достаточное для того, чтобы выделить вещество, ответственное, как считалось, за перенос специфического навыка. Им оказался пептид, состоящий из 15 а.о. Он был назван скотофобином.
Впоследствии были выделены и другие пептиды, переносящие, как думали, определенные типы выработанных навыков. Однако детальные исследования показали, что получить чистые препараты, способные переносить закрепленные навыки, довольно затруднительно. Чем тщательнее производилась очистка выделяемой фракции, тем в меньшей степени осуществлялся перенос. Синтетический скотофобин, хотя в какой-то степени и вызывал у доноров боязнь темноты, но обладал значительно меньшей эффективностью, чем препараты, выделенные из мозга.
Методика экспериментов Г. Унгара была подвергнута многосторонней критике. Утверждалось, например, что животные во время опыта находятся в состоянии выраженного стресса и поэтому выделяемый фактор является агентом переноса не памяти, а стрессового состояния. Кроме того, результаты экспериментов, проведенных в разных лабораториях, в большой мере варьировали в зависимости от сравнительно небольших различий в особенностях применяемых методик. После многолетних исследований удалось установить, что факторы переноса навыков Г.Унгара, по всей вероятности, если и существуют, то не переносят непосредственно уже сформировавшийся навык, а лишь усиливают способность к его выработке при обучении реципиентов. На этом и основано мнение многих исследователей, что все факторы переноса на самом деле являются просто стимуляторами памяти, подобными уже упоминавшимся АКТГ и вазопрессину.
Таким образом, пока нет вполне строгих доказательств переноса энграммы с помощью специфических химических соединений, вырабатывающихся в мозге донора. Трудности экспериментального подтверждения переноса памяти связаны, в частности, с тем, что при выработке сложных поведенческих реакций может выделяться одновременно несколько факторов, каждый из которых обусловливает определенный компонент целостной реакции. Факторы эти могут взаимодействовать между собой достаточно сложным образом, усиливая или подавляя друг друга. В результате малейшие отклонения в экспериментальной процедуре могут приводить к неоднозначным результатам.
Говоря о механизмах химического переноса памяти, можно представить себе два варианта. Либо информация закодирована в структуре молекул переносимого вещества, либо эти вещества запускают в мозге реципиента функциональные системы, аналогичные тем, которые были активированы у донора. Имеющиеся факты не дают каких-либо свидетельств в пользу первого предположения. Судя по существующим данным, веществами-переносчиками, по всей вероятности, являются малые и средние по размерам пептиды. Однако трудно себе представить способность таких пептидов к переносу больших порций информации, усваиваемой при выработке того или иного навыка. К тому же до сих пор не установлена с достаточной определенностью структура таких пептидов.
Второй вариант, предусматривающий генетически обусловленное существование в мозге многочисленных схем обучения и их активацию определенными химическими агентами, представляется более вероятным. Одним из возможных механизмов такого переноса могут быть упоминавшиеся иммунологические процессы, обеспечивающие закрепление памяти. В свете гипотезы об иммунохимических механизмах памяти фактором переноса может быть либо соединение, вырабатывающееся в синапсе в результате приходящей к нему интенсивной импульсации и служащее антигеном, либо антителоподобные соединения, вырабатывающиеся в ответ на этот антиген. По химической природе эти факторы могут быть пептидами относительно малого размера.
Известно, что в целом ряде обычных иммунологических процессов антигенная детерминанта, т.е. участок молекулы антигена, ответственный за его специфичность, может представлять собой пептид с относительно небольшим количеством аминокислотных остатков. Тем более, что возможность существования антител в виде молекул, значительно меньших обычных иммуноглобулинов, допустима в изолированной иммунной системе мозга. Здесь не требуется крупных белковых молекул, в которые, помимо узнающего участка, входит большой фрагмент для обеспечения изоляции и уничтожения чужого антигена. Для реализации процессов, связанных с памятью, может оказаться достаточным пептид, близкий по размеру к узнающему участку.
В результате таких иммунохимических процессов в мозге могут образовываться антигены или антителоподобные соединения. При попадании в мозг реципиента такие вещества способны найти синапсы или клетки глии, аналогичные тем, на которых они были синтезированы и, модифицируя их деятельность, обеспечить запуск соответствующей функциональной системы.
Таковы существующие представления о возможности переноса, или транспорта, запечатленной в мозге информации с помощью химических агентов. Окончательное же решение этого вопроса будет зависеть от дальнейших исследований, связанных с выделением, анализом структуры, синтезом и лабораторными испытаниями этих веществ.
Суммируя изложенные представления о молекулярных механизмах памяти, можно сделать заключение, что основой нейрологической памяти является изменение проводимости в сети синапсов после многократного повторения подходящих к синоптическим терминалям импульсов. Поэтому события на молекулярном уровне, непосредственно связанные с запоминанием, разыгрываются в значительной мере в синапсе, но с обязательным привлечением других биохимических систем нейрона и, по всей вероятности, клеток глии.
Следует, однако, иметь в виду, что такие представления находятся, по мнению ряда исследователей, в противоречии сданными о возможности "научения" отдельного нейрона. В опытах с хорошо наблюдаемыми и всесторонне охарактеризованными нейронами моллюсков установлена возможность возникновения реакции нейрона на условный раздражитель. Раздражителем может служить, в частности, какое-либо вещество, подводимое к нейрону. После ряда одновременных воздействий этого вещества и фактора, который при всех условиях вызывает реакцию нейрона, оказывается возможным выработать реакцию клетки только на избранное вещество.
Пока еще трудно судить о биохимических механизмах этого явления. Однако попытаемся ответить на вопрос, каково его возможное значение в механизмах запоминания целого мозга.
Рассмотрим сначала это явление с позиций филогенеза. Развитие у одноклеточных организмов каких-то форм привыкания, сенситизации и запоминания, кроме генетической памяти, охарактеризованной выше, не встречает каких-либо принципиальных возражений. Нет и априорных возражений против того, что при объединении единичных клеток по ходу эволюции в многоклеточные организмы индивидуальные механизмы клеточной памяти не утрачивались, а вступали в какие-то отношения с другими системами. Однако можно ли, опираясь на достигнутый уровень знаний, сравнивать относительную значимость для многоклеточных, особенно высших, животных памяти огромных нейрональных ансамблей, с одной стороны, и простую сумму памяти индивидуальных нейронов, с другой?
Выше уже отмечалось соответствие реально достижимой информационной емкости нейрологической памяти с характеристиками численности и взаимосвязанности нейронов мозга. Определена и важнейшая роль состояния мозга для формирования не только кратковременной, но и долговременной памяти. В мозге высших животных и человека отмечено наличие отделов, зон и т.д., которые в разное время и по-разному участвуют в процессе консолидации памяти. Хотя память следует рассматривать как функцию всего мозга, это не значит, что не существует сложной системы "разделения труда" между отдельными нейронами и их группами. Все это является свидетельством качественно нового уровня, достигнутого в филогенезе после перехода к многоклеточным организмам. Поэтому можно полагать, что способность отдельного нейрона к простейшим формам обучения и запоминания каким-то образом интегрирована в высших полинейрональных синаптических механизмах. Изучение механизмов этой интеграции сейчас только начинается. Однако уже ясно, что нет оснований для противопоставления синаптических и "мононейрональных" гипотез о природе нейрологической памяти.
Различные по продолжительности и по ряду других признаков формы памяти обеспечиваются качественно различными механизмами, которые, однако, тесно связаны друг с другом. На первом этапе основными факторами являются локальные и кратковременные изменения концентраций калия и кальция в непосредственной близости от синаптических мембран. При более продолжительной форме памяти в процесс могут быть вовлечены другие механизмы, которые первоначально запускаются, с одной стороны, изменениями потоков и концентраций указанных ионов, а с другой — прямым действием медиаторов, а также нейропептидов-спутников. К таким процессам, в первую очередь,* относится изменение степени фосфорилирования белков мембран, происходящее за счет кальций- или циклонуклеотид-зависимых реакций. При этом происходит протеолиз и модификация синтеза некоторых мембранных белков. Совокупность всех этих процессов может определять ООП в различных ее формах, имеющих разную продолжительность — от нескольких секунд до нескольких суток. При этом надо иметь в виду, что нейрохимические перестройки могут затрагивать не только синаптический аппарат, но и изменять деятельность самого нейрона. Предполагается, что помимо соединений, изменяющих функционирование синапса в мозге, существуют специальные информационные вещества, действующие в составе так называемой парасинаптической системы, т.е. минуя синапс.
В этот же период происходят процессы консолидации, формирующие наиболее продолжительную, пожизненную форму памяти — ДПП. При этом включается или усиливается синтез некоторых белков, возможно тех же, которые определяют длительную форму ООП. Эти белки могут либо индуцировать перестройку регуляторной системы в геноме нейрона, так что обеспечивается поддержание измененного состава синаптических мембран в течение всей жизни, либо включать иммунохимические механизмы в глиальных клетках, обусловливающие постоянное воздействие на состояние синапса путем выработки антителоподобных соединений, специфически связывающих белки синаптической мембраны данного нейрона или группы нейронов.
В целом, развитие представлений о молекулярных механизмах памяти идет от попыток создания универсальных гипотез, стремящихся объяснить все процессы, связанные с запоминанием в целом, к разработке все более усложняющихся представлений о большой и разветвленной цепи реакций, определяющих различные формы нейрологической памяти. В связи с этим следует обратить внимание на одно важное обстоятельство. Имеющиеся в настоящее время данные заставляют предполагать, что формы нейрологической памяти и обусловливающие их нейрохимические механизмы могут определяться не только временными характеристиками, но и тем, какая именно информация запоминается или какой тип условной реакции вырабатывается. Например, ингибиторы кальций-зависимых протеиназ, о роли которых в формировании нейронной пластичности и ООП уже говорилось, могут нарушать выработку только отдельных видов условных реакций, оставляя интактными другие. Аналогичные факты известны относительно некоторых пептидных регуляторов памяти и веществ, влияющих на классические медиаторы. Существуют типы поведенческих реакций, формирование которых стимулируется вазопрессином и не затрагивается АКТГ, и наоборот. В некоторых случаях норадре-налин стимулирует формирование одних форм поведения и ухудшает приобретение других, а серотонин действует противоположным образом. Имеются и другие примеры, указывающие на то, что нейрохимические механизмы, ответственные за формирование памяти, могут быть различными в зависимости от содержания запоминаемой информации.
Таким образом, к настоящему времени не существует единой непротиворечивой теории, способной объяснять всю совокупность фактов, связанных с природой памяти. Проблема эта является чрезвычайно сложной и потребует еще своего переосмысления с позиций различных подходов. Однако несомненный прогресс, наметившийся в последние годы в этой области, и широкий фронт проводящихся исследований позволяет ожидать в ближайшие годы существенных качественных сдвигов в расшифровке этих механизмов.
Выводы
1. Нейрологическая память обладает сложной системной организацией и не имеет строгой локализации в определенных участках мозга. По современным представлениям, следы памяти фиксируются в мозге в виде изменений состояния синаптического аппарата, в результате которых возникает предпочтительное проведение возбуждения по определенным нервным путям.
2. После восприятия информации, в процессе ее запечатления и фиксации, в мозге протекает ряд последовательно сменяющихся нейрохимических процессов. На первых этапах, в стадии минимальной кратковременной памяти продолжительностью не более секунды, происходят изменения "быстрых" функций синапса, связанные с выбросом и сдвигом концентрации "классических" и пептидных медиаторов. В дальнейшем, в фазе относительно ограниченной во времени памяти, длящейся от нескольких секунд до нескольких суток, первоначальные синаптические процессы, связанные с изменениями концентрации ионов кальция или циклазными системами, могут вовлекать широкий спектр нейрохимических процессов, включающих изменения в составе и структуре нейроспецифических белков, в частности изменения степени их фосфорилирования, а также модификацию синтеза РНК.
3. Для формирования пожизненной долговременной памяти необходим постоянный синтез новых биополимеров, который может быть осуществлен в случае устойчивых перестроек в функционировании участков генома. Последние могут происходить в результате либо структурных изменений ДНК, либо образования устойчивых циклов для постоянного синтеза репрессоров или дерепрессоров транскриггтонов. Возможно также, что в формировании долговременной памяти принимают участие иммунологические механизмы, благодаря которым в мозге синтезируются антителоподобные соединения, способные в течение длительного времени модифицировать деятельность синапсов в определенных нервных путях.
4. В механизмах формирования памяти принимают участие как "классические" медиаторы, так и большое число нейропептидов, выполняющих функции медиаторов и нейромодуляторов. Среди таких нейропептидов наиболее изучены вазопрессин и фрагменты АКТГ, введение которых в организм в небольших дозах значительно стимулирует процессы, связанные с запоминанием и извлечением информации из памяти. Существенно при этом, что влияние пептидных гормонов на память не связано с их собственно гормональными функциями.
5. Имеются сведения о том, что при обучении в мозге животных вырабатываются определенные олигопептиды, которые способны при введении необученным животным стимулировать у них выработку аналогичного навыка. Однако конкретные механизмы такого "транспорта памяти" пока не известны.
Похожие работы
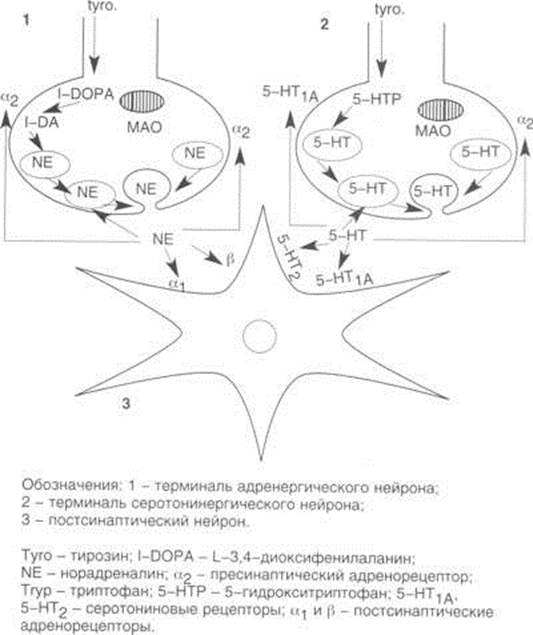
... уровне взаимодействия норадренергических и серотонинергических нервных окончаний с постсинаптическим нейроном. Общая характеристика и классификация антидепрессантов Современная классификация антидепрессантов, основанная на особенностях механизма их действия, фармакологического и нейрохимического профиля, принятая в экспериментальной и клинической психофармакологии, представлена в табл. 1. ...
... информации. Отбор информации осуществляется благодаря реакции внима- ния,рассмотренной ранее.Остановимся теперь более подробно на механизмах формирования энграммы. Важный вклад в создание современной теории памяти внес канадский физиолог Хебб.Он предположил,что внешний стимул сразу приводит к образованию лабильного следа памяти в моз- ге,который вскоре исчезает.Переход информации в долговремен- ...
... Хиден высказал предположение, что именно молекула РНК является главным нейрохимическим субстратом памяти. Опыты по изучению активности головного мозга в процессах запоминания и воспроизведения. Ключи к разгадке феномена памяти — в активности нашего головного мозга. Запоминание и узнавание уже знакомых объектов осуществляется задней и передней областями коры головного мозга. Человек обладает ...
... перекрест рук» А.Р. Лурия в общей структуре латеральных профилей мужчин и женщин в норме. Обоснованы принципы подхода к проблеме связи вариантов латеральных профилей с индивидуальными различиями с позиций дифференциальной психофизиологии, приведены нейрофизиологические и нейрохимические объяснения выявленным связям между индивидуальными профилями латеральности и некоторыми особенностями реализации ...
0 комментариев