Навигация
2.2.4 ОСНОВНЫЕ ТИПЫ ЩЕТИНОК
Различают две категории «дефинитивных щетинок; простые — состоящие лишь из одной части, и сложные — образованные из двух частей. Последние имеют более длинный основной ствол (или рукоятку, иначе древко) и сочлененный с ним дистальный членик. Сочленение подвижное и при упоре щетинки в субстрат его членик своим зазубренным краем ложится параллельно субстрату, в результате чего усиливается сцепление. Сложные щетинки среди рассматриваемых нами семейств имеются лишь у Sigalionidae и Chrysopetalidae. Дистальный членик щетинок сигалионид иногда сильно вытянутый, со вторичными суставами и двузубым кончиком (рис. 10, 13). Сложные щетинки с коротким члеником, напоминающим серп, обозначаются как серповидные, а с удлиненным, напоминающим косу, — косовидные. Сложные щетинки присущи только невроподии, выполняющей основную локомоторную функцию.
Некоторые типы простых щетинок характерны для вполне определенных родов и имеют большое систематическое значение. Так, крупные игловидные нотоподиальные щетинки темного цвета свойственны Aphrodita. Такие же щетинки, но с загнутыми назад на их дистальном конце боковыми крючьями,
так называемые гарпунообразные (рис. 10, 1), весьма типичны для Hermonia и Laetmonice, причем для последнего рода не менее характерны также невро-лодиальные «гребенчатые» щетинки (рис. 1.0, 2).

Рис. 10. Основные типы щетинок Aphroditiformia. Polynoidae'.

Наиболее обычный тип нотоподиальных щетинок полиноид — слегка изогнутые разной толщины щетинки с многочисленными плотно расположенными поперечными рядами мелких зубчиков (рис. 10, 4, 5); дистальный их кончик нередко волосовидный (Gattyana, Arcteobia), в других случаях гладких закругленный или даже притуплений, иногда с расщепленной верхушкой.В невроподии наиболее обычны щетинки с загнутым однозубым или двузубым концом (рис. 10, 6, 7). У этих щетинок различают гладкую рукоятку, или стержень, и несколько расширенную зазубренную дистальную часть. На дистальной части зубчики расположены с двух сторон, охватывая веером основную' часть щетинки; боковые зубчики могут иметь сложную дополнительную скульптуру, которая обнаруживается на фотографиях сканирующим микроскопом (рис. 11). Длина зазубренной части и количество поперечных рядов зубчиков на ней могут служить видовым признаком.

Рис. 1 Рис.11. Боковые зубчики на дистальной части щетинки Perolepis. (Фото сканирующим микроскопом В. Г. Аверинцева).
Количество щетинок в одном пучке как в нотоподии, так и в невроподии достигает 50 и более. Верхние и нижние щетинки в одном пучке несколько различаются.
По мере износа в параподиях появляются новые тождественные щетинки. Каждая щетинка формируется одной базальной клеткой, или хетобластом, расположенным на дне так называемого щетинкового мешка эпидермального происхождения. На апикальной поверхности хетобласта имеется сложно устроенный аппарат микроворсинок (microvilli), который и определяет структуру секретируемого хетобластом вещества будущей щетинки. Рост щетинок — базальный, и по мере их роста аппарат микроворсинок может перестраиваться, в результате чего меняются форма и характер самих щетинок. Как показали еще Щепотьев (1903, 1904) и Любищев (1924а), а затем и ряд других исследователей, ствол щетинок пронизан многочисленными каналами, оставшимися в результате деятельности микроворсинок, т. е. имеет как бы трубчатую структуру. По своему тонкому строению полихетные щетинки весьма схожи с таковыми погонофор, эхиурид, брахиопод и хетогнат, и эта ультраструктура не может быть использована для филогенетических построений.
Щетинки многощетинковых червей содержат хитин в разных пропорциях. У Aphrodita aculeata дорсальные крупные игловидные щетинки имеют хитина до 38% от сухого веса, а тонкие волосовидные латеральные и дорсальные щетинки только 19—20%. Хитин имеется также в щетинках эхиурид, погонофор и хетогнат. В других каких-либо морфологических структурах полихет, а также в их трубках хитин пока не отмечен; в частности, он не обнаружен в элитрах Aphrodita и в челюстях полиноид.
2.2.5 ЭЛИТРЫ И ИХ ЗАЩИТНАЯ РОЛЬ
Элитры, или спинные чешуйки, представляют собой видоизмененные спинные усики, сплющенные дорсовентрально. Элитры, так же как и спинные усики, сидят на специальных выступах стенки тела — элитрофорах, которые, однако, располагаются несколько выше, чем подставки спинных усиков— циррофоры на усиковых сегментах. Такое расположение элитрофор и циррофор находит свое объяснение в том, что элитры предназначены для прикрытия спины (защитная функция), а спинные усики — осязательные органы и, возможно, хеморецепторы, в соответствии с чем они должны быть выставлены по возможности дальше в сторону.
Попытка гомологизировать элитрофоры с «дорсальными бугорками» (dorsal tubercle), расположенными на усиковых сегментах поверх спинных усиков, оказалась несостоятельной. В онтогенезе элитры и спинные усики закладываются однотипно. В морфо-функциональном отношении элитры следует рассматривать как новообразующиеся органы, а потому вначале они имели неупорядочную закладку и лишь затем, в процессе эволюции, путем олигомеризации произошло их упорядочение и строго закономерное расположение на определенных сегментах тела.
У большинства чешуйчатых червей элитры почти полностью прикрывают спинную сторону, вдоль которой непрерывно спереди назад движется ток свежей воды, используемый червями в респираторных целях. Задняя часть каждой чешуйки в той или иной степени налегает на передний край следующей за ней элитры, т. е. они образуют как бы черепичную кровлю. Лишь у представителей Polyodontidae элитры располагаются в обратном направлении: задняя элитра находит на переднюю. Последнее у полиодонтид, возможно, связано с их постоянным обитанием в длинных трубках и необходимостью при нападении хищников быстро уходить задним концом тела в глубину своего домика, в то время как другие чешуйчатые черви преимущественно двигаются передним концом тела лишь вперед.
Элитры прикреплены к элитрофорам обычно центральной своей частью (место прикрепления округлое или слабоовальное). Однако бывает и боковое прикрепление. Последнее характерно для всех Aphroditidae, а среди полиноид наблюдается у многосегментных Perolepis, которых среди полиноид мы считаем наиболее примитивными формгми. Можно наметить постепенные переходы от латерально прикрепленных крохотных элитр Perolepis и небольших, едва достигающих ширины сегментов округлых элитр Lepidasthenia — к крупным жестким почковидным (бобовидным) элитрам Lepidonotus, полностью прикрывающим всю спинную сторону червей (рис. 12). Этот ряд наглядно доказывает происхождение элитр из цирростилей спинных усиков.
Форма и размеры элитр несколько изменяются в зависимости от части тела, в которой они находятся: как правило, передние элитры несколько меньшего размера и округлой формы, в то время как элитры в средней части тела наиболее вытянутые и наиболее крупные. Но имеются и исключения: у некоторых Perolepis и Lepidasthenia наиболее крупными, в несколько раз превышающими все остальные, являются элитры первой пары. В большинстве случаев элитры покрывают друг друга вдоль всей медиальной линии спины, однако у некоторых видов средняя часть спины остается неприкрытой.
У всех чешуйчатых червей в передней части тела до XXIII сегмента расположение элитр одинаковое: II, IV, V, VII, IX, XI, XIII, XV, XVII, Х1Х,: XXI и XXIII сегменты. Дальше чередование чешуйчатых и усиковых сегментов варьирует. Элитры нередко находятся через два усиковых сегмента, в других случаях — через один. Иногда элитры располагаются асимметрично, т. е. на одном и том же сегменте с одной стороны наблюдается элитра, а на другой стороне — спинной усик (Polyeunoa, Arctonoe). У большинства червей элитры прикрывают тело почти до самого заднего его конца. Однако у рода Polynoe значительная задняя часть тела свободна от элитр. Сигалиониды в задней части тела, начиная с XXIII (иногда с XXIX) сегмента, имеют элитры на каждом сегменте. Характер чередования чешуйчатых и усиковых сегментов в задней части тела используется как один из родовых признаков.
Морфология чешуек относительно простая. В элитрофор, представляющий собой простое выпячивание стенки тела, частично заходит целом. В своей ди-стальной части элитрофор системой мощных кольцевых мышц отделен от собственно элитры (своего рода сфинктер). Резкое сокращение этих мышц при раздражении червя обеспечивает аутотомию элитр. Сами элитры имеют разнообразную фактуру. Некоторые из них очень нежные, мягкие, другие же, напротив, жесткие, сильно склеротизированные.
В качестве крайнего примера жестких элитр можно привести элитры Iphione muricata. Элитры этого червя имеют бросающуюся в глаза ячеистую (полигональную) структуру, напоминающую собой пчелиные соты («elytra reticulata»). Почти вся их наружная поверхность сложена из крупных, хорошо отгороженных друг от друга шестигранных ячеек, которые в свою очередь имеют многочисленные более мелкие ячейки. Последние не что иное, как отдельные клетки с толстой клеточной мембраной. Внутри этих клеток имеется по крупному ядру, причем плазма прижата к стенкам клеток. Под первым (верхним) слоем наиболее крупных клеток располагаются второй, третий, четвертый и даже пятый слои из более мелких клеток.
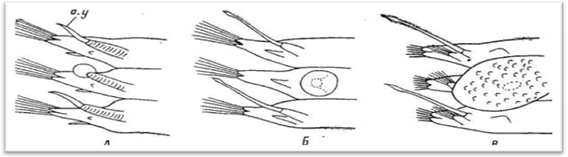
Рис.12 Изменение формы параподий и элитр в ряду Perolepis (А) — Lepidasthenia (Б) — Lepidonotus (В).
Все клетки плотно прижаты друг к другу, и вся эта конструкция напоминает растительную ткань. Крупные ячейки отделены друг от друг поперечными прослойками толстой кутикулы. Такого рода жесткие элитры, полностью прикрывающие спинную сторону червя, крепко прижавшегося к скалам, предохраняют его от подсыхания и позволяют ему в неподвижном состоянии находиться долгие часы под палящим тропическим солнцем. Ячеистая структура элитр, но менее жесткая, чем у Iphione muricata, наблюдается также у Gattyana deludens, G. iphionelloides, Harmothoe areolata, H. reticulata, H. dictyophora и некоторых других.
У мягких элитр, к которым относятся элитры H", lunulata и некоторых других полиноид, лишь верхняя и нижняя их поверхности имеют небольшой слой кутикулы, богатой кератиноидами. Изнутри кутикула выстлана одним слоем призматического эпителия, а все остальное внутреннее пространство элитры заполнено соединительной тканью, пронизанной единичными мышечными волокнами. Через ствол элитрофора в элитры проникают нервы, имеющие связь с брюшной нервной цепочкой. У места прикрепления элитры к элитрофору обычно наблюдается густое нервное сплетение (plexus), от которого отходят в разные стороны древовидно ветвящиеся небольшие нервные волокна, хорошо заметные при рассмотрении элитр в проходящем свете. Тончайшие нервные фибриллы заканчиваются, с одной стороны, в фотогенных клетках (фотоцитах), находящихся у нижней поверхности элитры, а с другой — в мягких папиллах, нередко густо покрывающих почти всю верхнюю сторону элитр, в особенности у их наружного края (на нижней стороне элитр папилл нет, эта поверхность всегда гладкая, глянцевидная). Мягкие папиллы, представляющие собой небольшие чувствительные органы, могут быть как одноклеточными образованиями, так и многоклеточными.
На верхней стороне элитр, помимо мягких папилл, обычно наблюдаются и многочисленные склеротизированные папиллы весьма разнообразной формы. Последние не иннервированы и, видимо, не выполняют чувствительных функций. Склеротизированные образования могут быть в виде небольших острых иголочек, крупных заостренных шипов или наподобие грибовидных образований с закругленной или со звездчатой верхушкой, иногда в виде бугорчатых бородавок, древовидно разветвленных кустиков и т. д. Микроструктура этих образований и их расположение имеют большое видовое таксономическое значение. Детали скульптуры верхней поверхности элитр хорошо обнаруживаются с помощью растрового электронного микроскопа.
Элитры некоторых полиноид (Polynoe scolopendrina, Acholoe astericola, различные Harmothoe, Lepidasthenia и др.) при раздражении светятся. Излучение света внутриклеточное (гранулы в апикальных частях фотогенных клеток). Свечение происходит в виде коротких и повторных вспышек и контролируется ганглиями брюшной нервной цепочки (рефлекторные реакции). При сильном раздражении одновременно аутотомируются и сами элитры, которые продолжают люминесцировать в оторванном состоянии достаточно долгое время. Последнее в некоторых случаях, возможно, вводит в заблуждение напавшего на полиноиду хищника, и пока оторванные элитры светятся, сам червь «обманным путем» ускользает от врага. После аутотомии элитр червь их регенерируют полностью. Недавно было констатировано, что элитры могут служить своеобразной выводковой камерой для молоди, наподобие крышечек некоторых Spirorbis. Так, в элитрах небольшой сигалиониды Pholoe swedmarki, адаптированной к интерстициальному биотопу на Бермудских островах, а также у Ph. antipoda Hartman, 1967 из антарктических вод (Аверинцев, in litt.) обнаружены вполне сформировавшиеся молодые черви, имеющие до 6 сегментов.
0 комментариев