Навигация
Плазмиды. Понятие и типы плазмид
17608
знаков
0
таблиц
0
изображений
2. Плазмиды. Понятие и типы плазмид
Наиболее распространённым методом генной инженерии является метод получения рекомбинантных (содержащих чужеродный ген) плазмид, которые представляют собой кольцевые, двухцепочечные молекулы ДНК, состоящие из нескольких пар нуклеотидов и способные к автономной репликации [4].
Для плазмид характерно стабильное существование в нехромосомном состоянии в бактериях. Каждая бактерия помимо основной, не покидающей клетку молекулы ДНК (5*106 пар нуклеотидов), может содержать несколько различных плазмид, которыми она обменивается с другими бактериями.
Плазмиды, размеры которых варьируют от нескольких тысяч до сотен тысяч пар оснований, а число копий на клетку - от одной до нескольких сотен, способны к автономной (независимой от основной хромосомы) репликации и стабильно наследуются в ряду клеточных поколений.
Хотя многие плазмиды дают клеткам-хозяевам ощутимые селективные преимущества (устойчивость к антибиотикам, тяжелым металлам и т.п.), большинство из них являются криптическими, то есть не проявляющимися в фенотипе клеток.
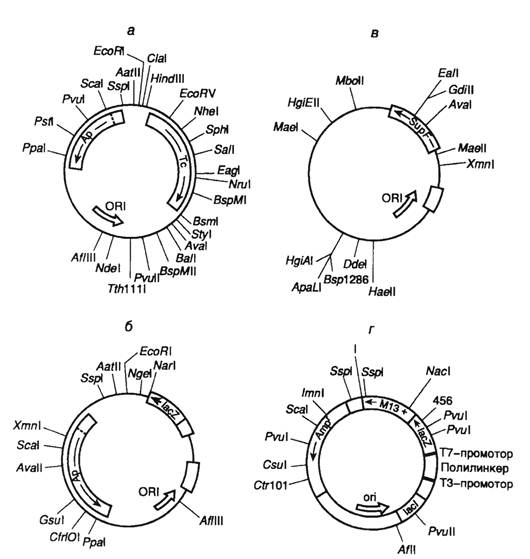
Область начала репликации небольшой плазмиды ColE1, несущей гены устойчивости к колицинам, традиционно используется в генной инженерии при конструировании векторных молекул ДНК, которые находят применение для клонирования и экспрессии в клетках E. coli коротких последовательностей нуклеотидов.
Плазмиды обнаружены у многих бактерий, принадлежащих к разным таксономическим группам. Количество плазмидной ДНК в клетке составляет обычно не более нескольких процентов от клеточного генома, а число плазмид колеблется от 1 до 38. Плазмиды - это линейные или кольцевые ковалентно замкнутые молекулы ДНК, содержащие от 1500 до 40000 пар нуклеотидов. Большинство плазмид состоит из трех групп генов: участка ДНК, ответственного за автономную репликацию плазмиды в клетке; системы генов, обеспечивающих возможность переноса плазмид из одной клетки в другую; генов, определяющих свойства, полезные для клетки-хозяина. Отличительная особенность плазмид - способность к автономной репликации, поэтому минимальное количество ДНК, которое может быть названо плазмидой, - это фрагмент, обеспечивающий автономную репликацию плазмидной ДНК в клетке как единого целого.
Обычно о присутствии плазмид в бактериальной клетке судят по проявлению определенных признаков, к которым относится устойчивость к отдельным лекарственным препаратам, способность к переносу генов при конъюгации, синтез веществ антибиотической природы, способность использовать некоторые сахара или обеспечивать деградацию ряда веществ.
Большинство бактериальных плазмид обладает способностью автономно реплицироваться, имеет фактор несовместимости и фактор переноса. Плазмиды несут множество специальных, детерминируемых каждой отдельной плазмидой маркеров: устойчивость к антибиотикам, тяжелым еталлам, ультрафиолетовому облучению, способность к биосинтезу токсинов.
В качестве векторов могут использоваться опухолеобразующие плазмиды бактерий. Виды Agrobacterium эволюционно родственны клубеньковым бактериям, относящимся к роду Rhizobium, и имеют много общих с ними черт. Однако характер взаимодействия агробактерий с растением имеет своеобразные особенности [6].
Взаимодействие видов Agrobacterium с растениями представляет особый интерес, так как при этом виде паразитизма один из партнеров специфически видоизменяет свойства хозяина, встраивая свои гены в его геном. Кроме того, это служит уникальным примером миграции ДНК прокариот в эукариотическую клетку. Хлоропласты и митохондрии содержат полноценную генетическую систему, то есть все компоненты, необходимые для экспрессии генетической информации: ДНК, ДНК-полимеразы, РНК-полимеразы и белоксинтезирующий аппарат (рибосомы, т-РНК, аминоацил-тРНК-синтетазы).
3. Получение плазмидНаиболее распространенным методом генной инженерии является метод получения рекомбинантных, то естьсодержащих чужеродный ген, плазмид.
Каждая бактерия помимо основной, не покидающей клетку молекулы ДНК (5-6 млн. пар нуклеотидов), может содержать несколько различных плазмид, которыми она обменивается с другими бактериями.
Плазмиды являются автономными генетическими элементами, реплицирующимися (то есть размножающимися) в бактериальной клетке не в то же время, что основная молекула ДНК. Хотя на долю плазмид приходится лишь небольшая часть клеточной ДНК, именно они несут такие жизненно важные для бактерии гены, как гены лекарственной устойчивости. Разные плазмиды содержат разные гены устойчивости к антибактериальным препаратам.
Плазмидные векторы, как правило, создают методом генной инженерии, так как природные (немодифицированные) плазмиды лишены ряда обязательных для «высококачественнного вектора» свойств:
- небольшого размера, так как эффективность переноса экзогенной ДНК в E.coli снижается при длине плазмиды более 15 тысяч пар нуклеотидов;
- наличие сайта рестрикции, в который осуществлена вставка;
- наличия одного или более селективных генетических мркеров для идентификации реципиентных клеток, несущих рекомбинантную ДНК.
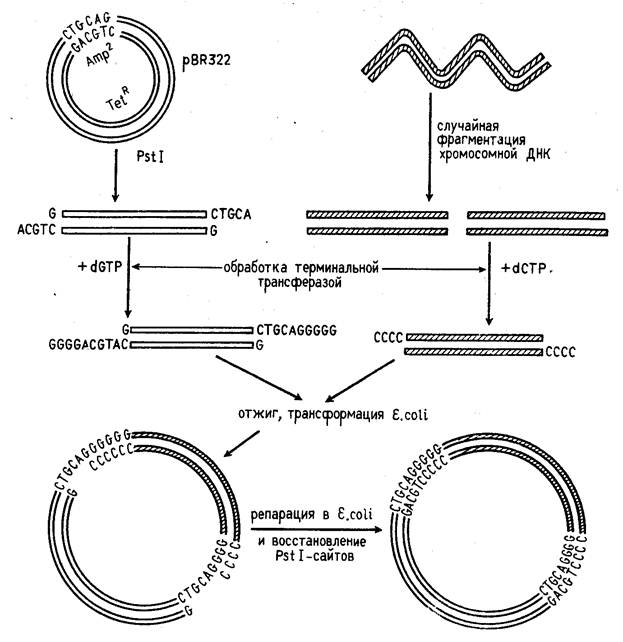
Для получения рекомбинантной плазмиды ДНК одной из плазмид расщепляется выбранной рестриктазой. Ген, который нужно ввести в бактериальную клетку, расщепляют из ДНК хромосом человека с помощью рестриктазы, поэтому его «липкие» концы являются комплементарными нуклеотидным последовательностям на концах плазмид.
Ферментом лигазой «склеивают» оба куска ДНК в результате получается рекомбинантная кольцевая плазмида, которую вводят в бактерию E. coli. Все потомки этой бактерии (клоны) содержат в плазмидах чужеродный ген. Весь этот процесс называют клонированием.
Вводят плазмиды в соматические клетки с помощью химических реагентов, повышающих проницаемость клеточной оболочки. В частности, чтобы обеспечить проникновение в клетки плазмидной ДНК, их обрабатывают ледяным раствором кальция хлорида, затем выдерживают при 42°С в течение 1,5 минут. Эта обработка приводит к локальному разрушению клеточной стенки. Максимальная частота трансформации -10-3, то есть на каждую тысячу клеток приходится одна трансформированная. Частота трансформации не бывает 100%-й, затем используют схемы отбора, позволяющие идентифицировать трансформированные клетки [2].
В качестве маркеров плазмида может содержать гены, определяющие устойчивость бактерии к антибиотикам. Вставка чужеродного (донорного) гена в маркерный ген приводит к инактивации последнего. Это позволяет отличить трансформированные клетки, получившие векторную плазмиду (утратившие устойчивость к антибиотику), от клеток, получивших рекомбинантную молекулу (сохранивших устойчивость к одному, но утративших устойчивость к другому антибиотику). Этот прием называется инактивацией маркера вставки.
Для отбора трансформированных клеток, содержащих рекомбинантную ДНК (гибридную плазмиду), проводят тестирование на резистентность к определенным антибиотикам. Например, клетки, несущие гибридную плазмиду, устойчивы в ампициллину, но чувствительны к тетрациклину (в маркерный ген которого и внедрена донорная ДНК).
Процесс разделения геномной ДНК на клонируемые элементы и введения этих элементов в клетки-хозяева называется созданием геномной библиотеки (банка клонов, банка генов).
Все системы клонирования должны отвечать двум основным требованиям:
1) наличию нескольких сайтов для клонирования;
2) возможности достаточно простой идентификации клеток с рекомбинантными ДНК.
Для всех рутинных процедур молекулярного клонирования широко используется E.coli в качестве клетки-хозяина. Клетки, способные поглощать чужеродную ДНК, называются компетентными; компетентность E.coli повышают, используя специальные условия культивирования. Для получения больших количеств чужеродных белков с помощью рекомбинантных штаммов E.coli была сконструирована плазмида, содержащая сильный промотор, селективный маркерный ген и короткий участок с несколькими уникальными сайтами для рестрицирующих ферментов – полиленкер.
Эффективными методами трансформации E.coli плазмидами является электропорация (воздействие на клеточные мембраны электрическим током для увеличения их проницаемости). Для введения клонированных генов в соматические клетки также применяют микроинъекции и микроукалывания или слияние с клеткой нагруженных ДНК мембранных везикул (липосом).
Заключение
Без преувеличения можно сказать, что прошлое, настоящее и будущее биотехнологии базируется на генетическом межвидовом и внутривидовом разнообразии организмов. Изменение уровня развития науки лишь способствует расширению и возникновению качественно новых способов использования этого разнообразия.
Так, интенсивное развитие фундаментальных исследований в биологии во второй половине ХХ столетия привело к существенному прогрессу в таких разделах биологии, как молекулярная биология и генетика, биохимия и энзимология, нейрофизиология и биофизика. Использование методологии точных наук (физики, химии, математики) позволило исследователям характеризовать различные жизненно важные процессы на уровне межмолекулярных взаимодействий. Расшифровка структуры ДНК и РНК, процесса реализации генетической информации привело к разработке так называемой ДНК-технологии, которая предоставила возможность исследователю работать с отдельными генами - конкретными участками генетического материала.
В результате появился высокоэффективный инструмент экспериментального изучения генетического материала: его организации, функционирования, взаимодействия различных элементов и эволюции. Для ряда наиболее генетически изученных организмов удалось получить, образно говоря, «чертежи» их генома.
Когда были разработаны методики выделения отдельных генов, подобраны условия сохранения их стабильности и определены закономерности переноса генов, появилась реальная возможность конструирования рекомбинантной ДНК (создание рекомбинантной ДНК буквально означает объединение (рекомбинирование) двух отрезков ДНК разных видов). По сути дела, наследственная изменчивость стала формироваться целенаправленно по воле и желанию человека, и были получены организмы, обладающие таким сочетанием генов (и соответственно, признаков), которые отсутствуют в природе.
В настоящее время для многих специалистов методы генной инженерии - методы рекомбинантных ДНК - являются краеугольным камнем здания биотехнологии.
Использование методов генной инженерии предполагает направленное, по заранее заданной программе конструировании молекулярных генетических систем вне организма с последующим введением их в живой организм.
Использование методологии генной инженерии в прикладном аспекте предполагает конструирование таких рекомбинантных молекул ДНК, которые при внедрении в генетический аппарат придавали бы организму свойства, полезные для человека.
Благодаря этому решаются многие прикладные проблемы: получение «биологических реакторов» - микроорганизмов, растений и животных, нарабатывающих фармакологически значимые для человека белковые препараты, создание высоко продуктивных пород животных с определёнными ценными для человека признаками, выведение растений, устойчивых к различным патогенам и вредителям и так далее. С этими же наукоёмкими технологиями связана и генетическая паспортизация, и диагностика генетических заболеваний, и создание ДНК-вакцин, и терапия различных заболеваний. Таким образом, сложившаяся благоприятная ситуация в биологии явилась мощным толчком в развитии современной биотехнологии, весьма важной области практического приложения результатов фундаментальных наук.
Список литературы
1. Атанасов А. Биотехнология в растениеводстве. Новосибирск: ИЦиГСО РАН, 1993. – 241 с.
2. Барановов В.С. Генная терапия – медицина XXI века // Соросовский образовательный журнал. № 3. 1999. С. 3 – 68.
3. Бекер М.Е., Лиепиньш Г.К., Райпулис Е.П. Биотехнология. М.: Агропромиздат, 1990. 334 с.
4. Глебов О.К. Генетическая трансформация соматических клеток // Методы культивирования клеток. Л.: Наука, 1988.
5. Глик Б., Пастернак Дж. Молекулярная биотехнология. Принципы и применение. М.: Мир, 2002.
6. Егоров Н.С., Самуилов В.Д. Современные методы создания промышленных штаммов микроорганизмов // Биотехнология. Кн. 2. М.: Высшая школа, 1988. 208 с.
7. Основы фармацевтической биотехнологии: Учебное пособие / Т.П. Прищеп, В.С. Чучалин, К.Л. Зайков, Л.К. Михалева. – Ростов-на-Дону.: Феникс; Томск: Издательство НТЛ, 2006.
8. Пирузян Э.С., Андрианов В.М. Плазмиды агробактерий и генная инженерия растений. М.: Наука, 1985. 280 с.
Похожие работы
... растений позволит значительно снизить её стоимость (Giddings et al., 2000). В заключение хотелось бы отметить, что несмотря на значительные достижения в области продукции реком-бинантных белков медицинского назначения в растениях, это направление находится лишь на начальном этапе своего развития. Учёные-биотехно-логи уверены, что в будущем рекомбинантные препараты, получаемые из генетически ...
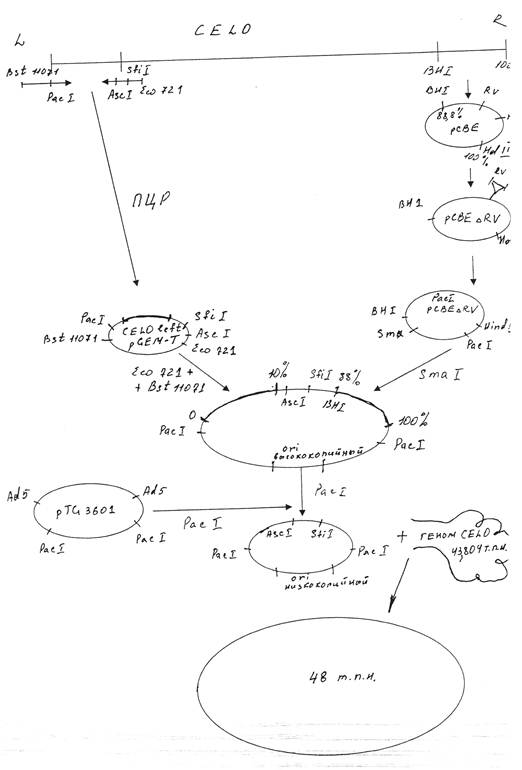
... в больших количествах (10 вирусных частиц из одного эмбриона), что значительно снижает стоимость получения препаративных количеств вируса. 2.2 Описание эксперимента Для получения рекомбинантного аденовируса CELO методом котрансформации с ДНК вируса CELO дикого типа сконструированная плазмида должна содержать правый и левый фрагменты ДНК указанного вируса. Мы использовали плазмиду pCBEl, ...
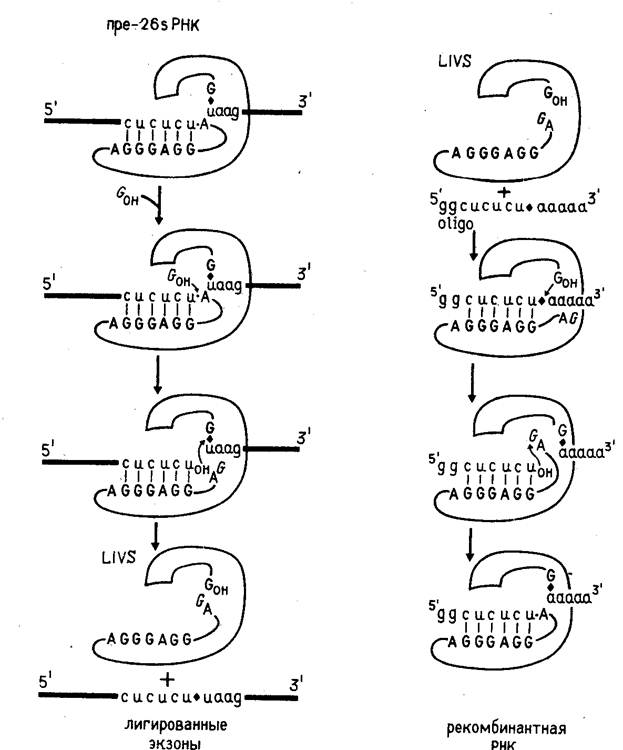
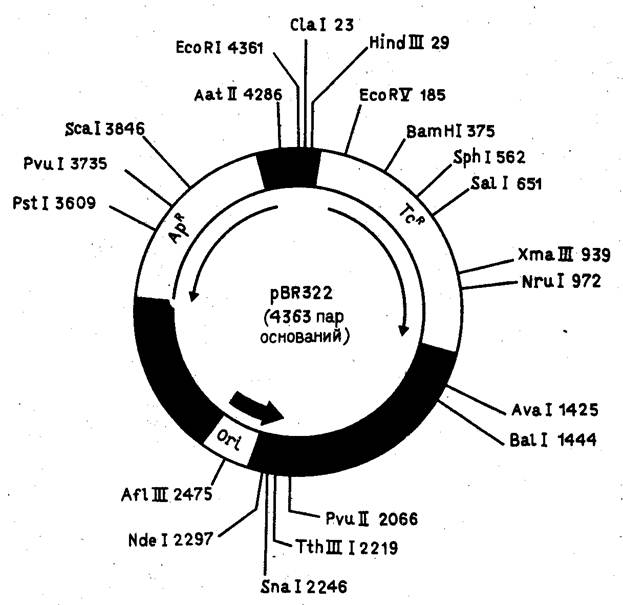
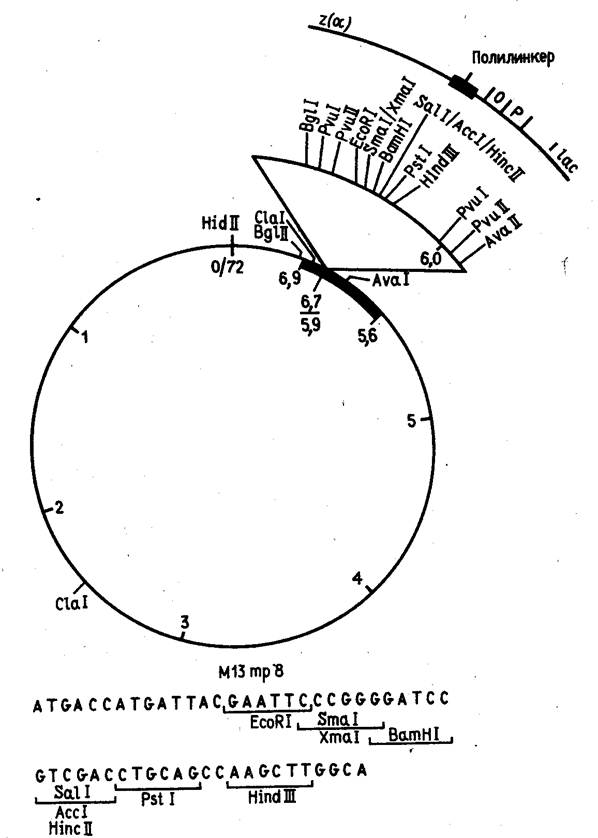
... для человека и животных. Именно на S. cerevisiae создан первый штамм-продуцент поверхностного антигена вируса гепатита Б, позволивший получить и испытать вакцину против данного вирусного заболевания человека. С появлением генной инженерии внимание многих исследователей привлекла система культивируемых клеток животных. Особый интерес к культурам клеток животных стал проявляться после обнаружения ...
... или путем достройки заглубленных концов с помощью ДНК-полимеразы I или обратной транскриптазы, а затем присоединяют линкеры. Инфекция, трансфекция и клонирование а. Перенос рекомбинантных молекул из пробирки в клетку Сконструированные рекомбинантные молекулы ДНК вводят в клетки или вирусные частицы для клонирования и амплификации. Для разных систем хозяин-вектор применяются разные методы. ...
0 комментариев