Навигация
Рекомбинантные вакцины (Генная инженерия)
43921
знак
0
таблиц
5
изображений
ОГЛАВЛЕНИЕ
Введение………………………………………………………………………………………….3
1. Принципы конструирования рекомбинантных противовирусных вакцин…………..4-24
1.1. Получение соответствующего фрагмента нуклеиновой кислоты……………....….4
1.2. Выбор высокоактивной и хорошо изученной в иммунологическом отно-
шении модели вектора-носителя и клонирование соответствующего гена……9-21
1.2.1. Получение рекомбинантных ДНК……………………………………………9
1.2.2. Получение рекомбинантных РНК…………………………………………...11
1.2.3. Стратегия клонирования генов………………………………………….…13
1.2.4. Разнообразие векторных молекул…………………………………………17
1.3. Выбор системы экспрессии клонированного гена, способной обеспечить
максимальный выход и функциональную полноценность продукта……………..22
1.4. Создание достаточно удобных и по возможности универсальных векторов
для целевой доставки генов в клетки и ткани организма…………………………23
2. Вакцина против лейкемии кошек, изготовленная с помощью генной
инженерии……………………………………………………………………………….…24
Список литературы…………………………………………………………………………….26
ВВЕДЕНИЕ
Существующие традиционные вакцины, несмотря на очевидный положительный эффект их широкого применения, обладают рядом недостатков. К ним относятся: наличие нежелательных биологически активных и балластных компонентов в препаратах, неполноценные иммунологические свойства самих антигенов. Кроме того, существуют заболевания, не вызывающие иммунитета, вакцины против которых вообще отсутствуют и не могут быть сконструированы на основе классических принципов. Все это вызывает необходимость усовершенствования уже существующих вакцин и создания принципиально новых типов вакцин. Одним из наиболее перспективных направлений в данной области является получение вакцинных препаратов на основе методов генной инженерии.
Последним достижением генной инженерии и биотехнологии стало создание рекомбинантных противовирусных вакцин, содержащих гибридные молекулы нуклеиновых кислот. Данные вакцины обладают целым рядом преимуществ. Они характеризуются отсутствием (или значительным снижением) балластных компонентов, полной безвредностью, низкой стоимостью, которая связана с удешевлением промышленного производства вакцин. Экспрессируемый в клетках вакцинированного животного белок имеет конформацию, близкую к нативной, и обладает высокой антигенной активностью.
Таким образом, рекомбинантные противовирусные вакцины являются новейшим поколением вакцин. Их очевидное преимущество обуславливает широкое применение данного типа вакцин в медицине и ветеринарии для вакцинации населения и сельскохозяйственных животных.
1. ПРИНЦИПЫ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНЫХ ПРОТИВОВИРУСНЫХ ВАКЦИН
Важным условием получения эффективного вакцинного препарата является соблюдение основных принципов его производства. Конструирование рекомбинантных противовирусных вакцин предполагает: 1) получение соответствующего фрагмента нуклеиновой кислоты; 2) выбор высокоактивной и хорошо изученной в иммунологическом отношении модели вектора-носителя и клонирование соответствующего гена (или генов); 3) выбор системы экспрессии клонированного гена, способной обеспечить максимальный выход и функциональную полноценность продукта; 4) создание достаточно удобных и по возможности универсальных векторов для целевой доставки генов в клетки и ткани организма.
1.1. ПОЛУЧЕНИЕ СООТВЕТСТВУЮЩЕГО ФРАГМЕНТА
НУКЛЕИНОВОЙ КИСЛОТЫ
Для конструирования рекомбинантных РНК или ДНК необходимо получить фрагмент нуклеиновой кислоты, который в дальнейшем и будет встраиваться в векторную молекулу.
Фрагменты ДНК для встраивания в вектор можно получить непосредственно из хромосомной ДНК, расщепив ее рестриктазами или разрушив случайным образом (например, с помощью ультразвука) на сегменты с примерно одинаковой длиной. Выделение генов с помощью "вырезания" из генома, как правило, состоит из четырех этапов: 1) получение клонотеки фрагментов генома; 2) выявление фрагментов генома, содержащих необходимый ген, и точная локализация гена в данном фрагменте; 3) вырезание гена из фрагмента(ов) с помощью рестриктаз и сшивка участков гена с помощью ДНК-лигазы фага Т4, если эти участки получены из различных фрагментов; 4) амплификация гена в составе векторной молекулы. Указанный способ получения генов является наиболее приемлимым применительно к протозойным возбудителям, бактериям и для некоторых сложно устроенных ДНК-содержащих вирусов. Такие операции проводятся, в частности, при создании так называемых "библиотек генов", то есть набора одинаковых векторных систем, в совокупности несущих в себе весь геном данного организма. Однако этот подход имеет ряд существенных недостатков. Во-первых, очень сложна задача подбора рестриктаз, позволяющих вырезать из геномной ДНК или клонированного фрагмента генома цельный ген. Как правило, вместе с геном остаются фланкирующие его лишние нуклеотидные последовательности, что мешает дальнейшему использованию этого гена, или же рестриктазы отрезают часть гена, делая его функционально неполноценным. Во-вторых, создание клонотеки генома возбудителя представляет специальную задачу и требует больших затрат времени. Наконец, для ряда ДНК-содержащих вирусов (паповавирусы) доказан сплайсированный характер структуры генов. Вполне понятно, что выделенные гены этих вирусов вследствие наличия интронных областей не будут проявлять функциональной активности в бактериальных клетках и окажутся непригодными при решении задач по конструированию рекомбинантных противовирусных вакцин. В-третьих, если ген составляет незначительную долю от всей геномной ДНК, то возникают большие трудности с его изоляцией и идентификацией.
Наиболее распространенным является путь получения генов через синтез ДНК-копий (кДНК) информационных или каких-либо других (в оптимальном случае - индивидуальных) РНК путем их обратной транскрипции. Данный способ включает в себя три этапа: 1) выделение высокоочищенных мРНК, кодирующих индивидуальные структурные белки, либо выделение геномных РНК вирусов; 2) синтез двухцепочечной ДНК (гена) на матрицах РНК; 3) амплификация гена с помощью методов молекулярного клонирования.
Как отмечалось, первым этапом в получении генов с помощью методов обратной транскрипции служит выделение и очистка геномных РНК и мРНК. Геномные РНК выделяют главным образом из очищенных вирионов, а мРНК - из инфицированных клеток. Для выделения мРНК из инфицированных клеток используют различные способы. В одном из вариантов выделение мРНК из инфицированных клеток проводят непосредственно из лизированных клеток (лизис осуществляют в денатурирующих средах в целях предотвращения разрушающего действия нуклеаз на мРНК) с последующей экстракцией фенолом и хлороформом или центрифигурированием лизата через подушку 5,7M СsCl (для освобождения от клеточных ДНК) и заключительной аффинной хроматографией на олиго(dТ)-целлюлозе (поли(У)-сефарозе). Выделение мРНК можно проводить и из очищенных полирибосом инфицированных клеток.
В случае необходимости получения специфических мРНК, кодирующих индивидуальные структурные белки возбудителя, используют следующий способ. Инфицированные вирусом клетки разрушают механическим путем или химическими методами (использование неионных детергентов). Клеточный лизат освобождают от ядер и митохондрий методом дифференциального центрифугирования и подвергают хроматографии на антительном сорбенте. При этой процедуре с антителами, специфичными к определенному структурному белку возбудителя, связываются полисомы, осуществляющие синтез этого белка. Также может быть использован метод непрямой иммунопреципитации полисом, при котором комплекс антитело - полисома преципитируется из раствора добавлением второго антитела, специфичного к первому. В качестве второго антитела чаще всего берут фиксированные формалином клетки Staphilococcus aureus, на поверхности которых находится так называемый белок А, имеющий сродство к Fc-фрагментам Ig. Из полисом, полученных одним из описанных способов, далее уже выделяют мРНК.
Другой путь выделения индивидуальных мРНК базируется на свойстве мРНК взаимодействовать с комплементарными ДНК, связанными с твердым носителем (целлюлоза, сефароза, нитроцеллюлозные фильтры). Этот метод отбора и очистки специфических мРНК является самым эффективным. Однако его применение возможно только при наличии соответствующих комплементарных ДНК.
Получение препарата очищенной мРНК позволяет перейти к работам по синтезу гена с помощью обратной транскрипции, которую осуществляют с помощью ревертазы AMV в специально подобранных условиях. В процессе обратной транскрипции матрицей служит мРНК, а затравкой олиго(dT12-18) (для мРНК, имеющих поли(А)-тракт) или химически синтезированный олигонуклеотид, комплементарный 3'-концу мРНК. После синтеза на мРНК комплементарной цепи ДНК и разрушения РНК (чаще используется обработка щелочью) осуществляют синтез второй цепи ДНК. В этой реакции матрицей служит первая цепь ДНК. Реакция может катализироваться как ревертазой, так и ДНК-полимеразой.
После получения двухцепочечной кДНК следует стадия получения гена, заключающаяся в гидролизе одноцепочечного участка ДНК, соединяющего первую и вторую цепи, нуклеазой S1.
Для получения необходимых участков РНК используют две группы способов: 1) расщепление полирибонуклеотидной цепи РНК в заданном месте; 2) синтез РНК (ферментативный на ДНК- или РНК-матрицах и химический).
Расщепление полирибонуклеотидной цепи РНК может осуществляться в присутствии различных ферментов. Для этой цели используют ферменты: рибонуклеазу H, нуклеазы, ковалентно связанные с олигонуклеотидами (например, стафилококковая нуклеаза), рибозимы (например, рибозим L-19).
Исторически первым способом направленной ферментативной фрагментации РНК (называемой также адресованной, сайт-направленной или сайт-специфической фрагментацией РНК) является разработанный в Московском университете метод гидролиза РНК ферментом РНКазой H в присутствии комплементарных олигодезоксирибонуклеотидов. Этот метод до сих пор остается наиболее универсальным способом направленной фрагментации РНК.
Метод основан на свойстве РНКазы Н расщеплять полирибонуклеотидную цепь РНК в составе ДНК-РНК-гетеродуплекса. К участку РНК, по которому планируется провести ее фрагментацию, синтезируется комплементарный олигодезоксирибонуклеотид длиной в 6-10 нуклеотидных остатков. Далее получают комплекс этого олигонуклеотида с РНК, который затем обрабатывают ферментом. В работе обычно используют РНКазу Н из Escherichia coli - вполне доступный фермент, который может быть довольно легко очищен от всех сопутствующих нуклеазных примесей. Этот метод нашел достаточно широкое применение для сайт-специфического расщепления вирусных и рибосомных РНК, а также для идентификации продуктов процессинга некоторых РНК.
Важным достоинством рассматриваемого метода является и то, что в результате гидролиза РНК рибонуклеазой Н образуются фрагменты, у одного из которых на 5'-конце содержится фосфатная группа, а у другого 3'-конец свободен (нефосфорилирован). Такие фрагменты можно прямо использовать в реакции ферментативного лигирования.
Серьезным ограничением этого метода является то, что участок РНК, с которым связывается комплементарный ему олигодезоксирибонуклеотид, должен иметь однотяжевую конформацию и находиться на поверхности макромолекулы РНК, представляющей собой в условиях расщепления компактную глобулу с развитой вторичной структурой. В отдельных случаях это ограничение удается преодолеть, используя достаточно длинные олигодезоксирибонуклеотиды (15-20-членные), которые предварительно отжигают с частично или полностью денатурированной РНК.
Другое ограничение метода фрагментации полирибонуклеотидов РНКазой Н в присутствии комплементарных олигодезоксирибонуклеотидов заключается в том, что в общем случае предсказать, какая из фосфодиэфирных связей в гетеродуплексе (или в непосредственной близости от него) подвергнется расщеплению, не удается. Более того, фермент зачастую гидролизует не одну, а несколько соседних межнуклеотидных связей. Ясно, что для последующего конструирования рекомбинантных РНК такие фрагменты могут оказаться непригодными.
Расщепление РНК может проводиться и с участием нуклеаз, ковалентно связанных с олигонуклеотидами. Идея, лежащая в основе данного подхода к направленной фрагментации РНК, восходит к концу 60-х годов, когда Н. И. Гриневой и ее сотрудниками был предложен метод олигонуклеотид-направляемой модификации нуклеиновых кислот. Принцип этого метода заключается в том, что с 5'- или 3'-концевым остатком олигонуклеотида, комплементарного заданному району ДНК и РНК, связывают модифицирующий агент, который после образования дуплекса атакует одно из ближайших к нему оснований. Реагентами этого типа удалось направленно фрагментировать фенилаланиновую тРНК из дрожжей, РНК-компонент (M1 РНК) РНКазы Р, а также 16S рибосомную РНК E.coli.
Однако, как и в случае РНКазы Н, расщепление РНК олигонуклеотид-нуклеазой зачастую проходит по нескольким межнуклеотидным связям, что несколько ограничивает возможности применения этого метода для получения рекомбинантных РНК.
Для расщепления РНК применяют также рибозимы (природные РНК и синтетические полирибонуклеотиды, способные катализировать целый ряд превращений у других РНК). Первый рибозим, напоминающий по своим свойствам эндонуклеазы рестрикции, был получен Т. Чеком. В научной литературе его обозначают как рибозим L-19. Этот рибозим представляет собой РНК длиной в 395 нуклеотидных остатков, в 5'-концевой области которой имеется гексануклеотидная последовательность GGAGGG, ответственная за специфичность расщепления предшественника 26S РНК при самосплайсинге. Эта последовательность комплементарна CUCUCU последовательности, расположенной на 3'-конце первого экзонного участка предшественника 26S РНК. Если эту РНК заменить другой, но обязательно содержащей доступную для комплементарного связывания CUCUCUA последовательность, то рибозим L-19 в присутствии гуанозина или гуаниловых нуклеотидов со свободной 3'-гидроксильной группой расщепит ее.
Источником необходимых участков РНК может служить такой способ, как синтез фрагментов РНК. Ферментативный синтез сегментов РНК осуществляют с использованием разнообразных генно-инженерных конструкций. Однако в настоящее время в подавляющем большинстве случаев для препаративного получения РНК используются РНК-полимеразы, закодированные в геномах ряда ДНК-содержащих бактериофагов (Т3, Т7 и SP6). Они характеризуются очень высокой активностью и, в отличие от клеточных РНК-полимераз, состоят из одной полипептидной цепи. Важно также то, что инициация и терминация синтеза РНК этими полимеразами происходит на одном определенном нуклеотидном остатке.
Полученные одним из способов фрагменты нуклеиновых кислот в дальнейшем встраивают в векторные молекулы.
Подводя итог изложенному, можно сделать вывод, что получение генов протективных белков возбудителей представляет собой достаточно сложную задачу. Однако ее решение необходимо для создания в конечном счете рекомбинантных противовирусных вакцинных препаратов.
Похожие работы
... олигонуклеотидов—одну полуавтоматическую, а вторую в комплексе с компьютером. В 1982 г. цена этих приборов на американском рынке составляла 36000—39500 долл.[2]. К открытиям связанным с достижениями генной инженерии нужно прибавить то, что огромный генетический «чертеж» многоклеточного существа просчитан полностью. Я думаю это можно назвать достижением века. После восьми лет работы многих ...
... генно-инженерных исследований. Многие из этих вопросов были подняты самими учеными активно работающих в данной области. В настоящее время большинство исследователей считали, что опасения касающиеся, генной инженерии, не имеют достаточно оснований, но многие этические проблемы остаются нерешенными и продолжают возникать новые. В прошлом генетика и медицинская генетика развивалась как относительно ...

... при постановке биологических или медицинских экспериментов. Составлять его должны ведущие специалисты в этой области и знающие предмет юристы. 3. Другие применения генной инженерии Несмотря на успехи ученых в генной инженерии в таких отраслях, как создание генно-модифицированных продуктов и клонировании животных и человека, они на этом не останавливаются. В целях сохранения военного ...
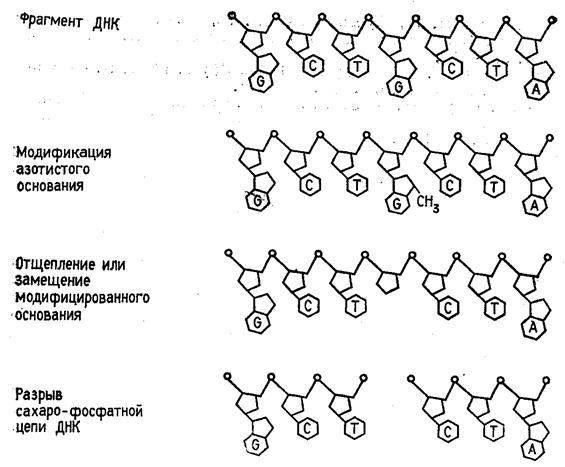
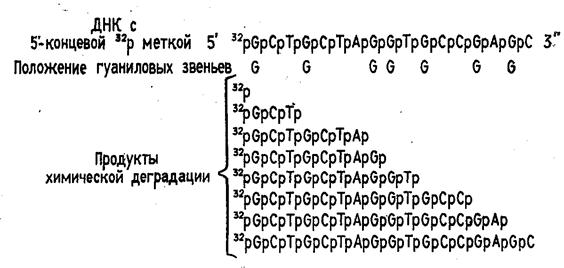
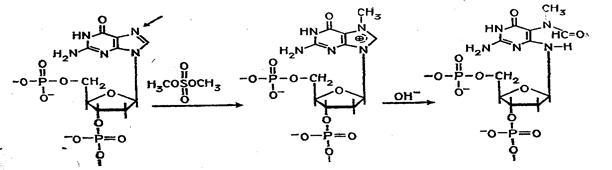
... (например, семейства генов) необходимо провести клонирование ПЦР-продуктов в векторах типа М13, в результате каждый фаг будет содержать только одну вставку. При прямом секвенировании смеси генов наблюдается несколько одинаково расположенных полос в разных дорожках геля. При амплификации же одного гена можно проводить прямое секвенирование, не прибегая к промежуточному субклонированию. Выбор ...
0 комментариев