Навигация
Охарактеризовать функции и строение корневых волосков
29007
знаков
0
таблиц
6
изображений
3. Охарактеризовать функции и строение корневых волосков
Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Поступление почвенного раствора в корень происходит преимущественно через зону всасывания, поэтому чем больше поверхность этого участка корня, тем лучше он выполняет свою основную всасывающую функцию. Именно в связи с этой функцией часть клеток кожицы вытянута в корневые волоски длиной 0,1—8 мм. Корневые волоски появляются в виде небольших сосочков — выростов клеток эпиблемы. Рост волоска осуществляется у его верхушки. Оболочка корневого волоска растягивается быстро. По прошествии определенного времени корневой волосок отмирает. Продолжительность его жизни не превышает 10—20 дней.
Почти всю клетку корневого волоска занимает вакуоль, окруженная тонким слоем цитоплазмы. Ядро располагается в цитоплазме возле верхушки волоска. Корневые волоски способны охватывать частички почвы, как будто срастаются с ними, что облегчает поглощение из почвы воды и минеральных веществ. Поглощению способствует также выделение корневыми волосками различных кислот (угольной, яблочной, лимонной, щавелевой), которые растворяют частички почвы.
Формируются корневые волоски очень быстро (у молодых сеянцев яблони за 30—40 ч). На 1 кв. мм корня при благоприятных условиях образуется до 300—400 корневых волосков, которые создают огромную поглощающую поверхность. У одной особи четырехмесячного растения ржи примерно 14 млрд. корневых волосков с площадью поглощения около 400 м2 и суммарной длиной более 10 тыс. км; поверхность всей корневой системы, включая корневые волоски, составляет примерно 640 м2, т.е. в 130 раз больше, чем у побега. Функционируют корневые волоски недолго — обычно 10—20 дней. Средняя медленность жизни волоска у винограда — от 10 до 40 суток. Сменяют отмершие корневые волоски в более нижней части корня новые. Таким образом, наиболее деятельная, всасывающая зона корней все время перемещается вглубь и в стороны вслед за растущими кончиками разветвлений корневой системы. При этом общая всасывающая поверхность корней все время увеличивается.
Каждый волосок представляет из себя вытянутую клеточку. В связи с ростом корня, перемещением и обновлением поглощающей зоны происходит константа смена волосков, обеспечивающая непрерывную активную работу корневой системы (Андросов, 2006).
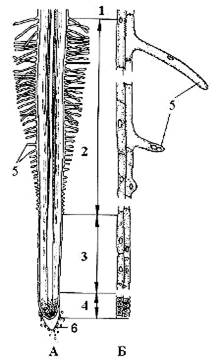
Рис. 1. Строение корня проростка пшеницы: А - схема строения корня; Б - дифференциация клеток ризодермы и экзодермы. 1 - зона проведения, 2 - зона всасывания, 3 - зона растяжения, 4 - зона деления, 5 - корневой волосок, 6 - корневой чехлик (Андросов, 2006).
4. В чем отличие анатомического строения однолетней и многолетней ветки одного и того же растения?
Анатомическое строение типичного стебля соответствует его главным функциям. В нем развита система проводящих тканей, часто очень сложная, которая связывает воедино все органы растения; с помощью механических тканей стебель поддерживает все надземные органы и выносит листья в благоприятные условия освещения.
Стебель, как и весь побег в целом, представляет собой «открытую» систему роста, т.е. он длительное время нарастает и на нем возникают новые органы. В стебле имеется система меристем, поддерживающих нарастание тканей в длину и толщину. Стебель существенно отличается от корня в том отношении, что формирование его тканей из верхушечных меристем не имеет той строго акропетальной последовательности, которая характерна для корня. Нарушение строго акропетального формирования тканей в стебле объясняется тем, что на апексе побега с правильной периодичностью возникают листовые примордии, что ведет к раннему вычленению узлов, а развитие междоузлий запаздывает.
Часто рост междоузлий и развитие в них постоянных тканей продолжаются длительное время за счет работы тех остаточных интеркалярных меристем, которые сохраняются у оснований нескольких междоузлий. Ярким примером такого интеркалярного (вставочного) роста может служить стебель злака, у которого апикальная меристема очень рано расходуется на образование соцветия, а быстрое вытягивание (колошение) обязано именно вставочному росту. Таким образом, для стеблей характерна сложная система меристем— верхушечных, боковых и вставочных.
В инициальном кольце под молодыми листовыми зачатками возникает прокамбий. Внутрь от прокамбия обозначается сердцевина, а наружу от него — первичная кора; самый поверхностный слой верхушечной меристемы образует протодерму (и в дальнейшем эпидерму).
В результате деятельности первичных меристем складывается первичная структура стебля. Если камбий работает слабо или совсем отсутствует, то она может сохраняться длительное время. Если камбий энергично откладывает вторичные проводящие ткани, то первичное строение быстро сменяется вторичным. Границу между ними иногда провести трудно, так как клетки камбия начинают делиться еще до окончательного формирования первичных тканей.
Камбиальный слой откладывает внутрь вторичную ксилему, а сам растягивается на ее поверхности и отодвигается от центра, оставаясь между ксилемой (древесиной) и корой. Одновременно камбий откладывает наружу слои вторичной флоэмы (луба). Первичная кора при длительном вторичном утолщении стебля погибает и заменяется вторичной корой, под которой понимают совокупность всех вторичных тканей, расположенных наружу от камбия. На поверхности стебля перидерма сменяет эпидерму. В результате всех этих преобразований складывается вторичное строение стебля (Билич, Крыжановский, 2005).

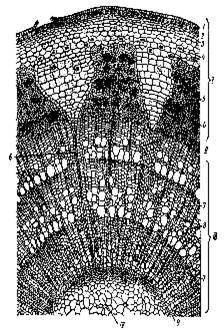
Рис. 2. Поперечный разрез молодого стебля липы: К-камбий; Колл-колленхима; Мкс- метаксилема; Мф-метафлоэма; П-паренхима; ПК- первичная кора; ПКс-первичная ксилема; Пркс- протоксилема; Прф- протофлоэма; ПФ-первичная флоэма; С- сердцевина; Эп- эпидерма. (Крамер, Козловский, 1983)
Рис. 3. Разрез трехлетней ветки липы: 1-кора, II-камбий, III-древесина, IV-сердцевина; 1- отмерший эпидермис, 2-перидерма, 3-колленхима, 4-клетки основной ткани с друзами, 5-участки флоэмы, 6-сердцевинный луч, 7-осенняя древесина, 8- весенняя древесина, 9-первичная ксилема. (Крамер, Козловский, 1983)
Развивающийся стебель непременно в той или иной мере подвергается утолщению. В зависимости от участия в этом процессе камбия утолщение может быть первичным или вторичным.
Первичное утолщение представляет собой увеличение объема стебля за счет растяжения клеток, составляющих первичные ткани. Этот процесс начинается уже в непосредственной близости от конуса нарастания и может осуществляться по-разному. Если преимущественно утолщается первичная кора, то утолщение называется кортикальным (лат. cortex - кора), что можно наблюдать у кактусов. Утолщение, которое осуществляется за счет сердцевины, называют медуллярным (лат. medullaris, medulla — костный мозг, мозговой), оно имеет место, в частности, при формировании картофельного клубня.
Вторичное утолщение приводит к увеличению диаметра стебля, что неизбежно ведет к разрыву и отмиранию первичной коры, на смену которой приходит кора вторичная, имеющая, совершенно иное строение. Она состоит из образовавшейся вместо эпидермы перидермы, остатков первичной коры и первичной флоэмы, а также вторичной флоэмы. Впоследствии в результате деятельности камбия, регулярно откладывающего вторичную флоэму, и феллогена, откладывающего пробку, формируется корка. У некоторых растений, например у пробкового дуба, образуется очень мощный слой корки, и человек широко использует ее в своей деятельности.
Вызванное деятельностью камбия вторичное утолщение продолжается в течение всей жизни растения и за много лет может привести к формированию ствола, имеющего колоссальный объем и диаметр, превышающий 10 м. У деревьев, растущих в дождевых тропических лесах, интенсивность деления камбиальных клеток очень мало изменяется в течение года, поэтому у них почти не выражена слоистость древесины. Напротив, у деревьев, произрастающих в условиях переменного климата, где сезонные колебания температуры и влажности заметны очень хорошо, уровень деятельности камбия находится в зависимости от времени года. Наибольшая интенсивность деления инициальных клеток здесь отмечается весной, когда из почек интенсивно развиваются молодые побеги, на которых имеются листья. В этот период в основном откладываются проводящие воду элементы — трахеиды (большая часть древесины образована именно ими) и членики сосудов, которые имеют большие внутренние объемы и тонкие оболочки. Летом инициальные клетки камбия делятся реже и дают начало толстостенным трахеидам и волокнам. У этих элементов функция проведения воды выражена крайне слабо или отсутствует совсем, зато они прекрасно выполняют функцию опоры. Осенью деятельность камбия ослабляется еще сильнее, а позднее прекращается вовсе, зимой камбий не функционирует. В результате образовавшаяся за год древесина неоднородна и ее можно подразделить на раннюю и позднюю, которые, чередуясь, совместно формируют ежегодный слой прироста в виде кольца.
Ширина и выраженность годичных слоев широко варьирует не только у представителей разных видов растений, она неодинакова даже у одного и того же экземпляра и напрямую зависит от влияния различных факторов окружающей среды. В годы с наиболее благоприятными погодными условиями (следует отметить, что этот термин для разных растений имеет совсем неодинаковый смысл - благоприятные погодные условия для одной экологической группы могут быть весьма неприятными для другой) слои прироста имеют наибольшую ширину, напротив, в неблагоприятные годы образуются узкие слои. Таким образом, пересчитав все годичные кольца, можно определить возраст дерева, измерив их толщину, иметь достаточно верное представление о метеорологических условиях давно прошедших лет, учитывая, что возраст наиболее старых деревьев может составлять не одно тысячелетие. Однако здесь необходимо соблюдать некоторую осторожность, потому что в особо неблагоприятных условиях годичные слои могут и не возникать. И наоборот, если летом у деревьев вредителями уничтожались листья, после некоторого покоя камбий откладывает слой древесины, по строению сходный с ранней весенней, что в итоге создает иллюзию двух годичных слоев.
Теперь становится понятным, что основная часть клеток многолетнего растения, способного к вторичному утолщению, состоит из мертвых клеток, которые сообща обеспечивают телу растения надежную опору (Матвеев, Тарабрин, 1989).
Похожие работы
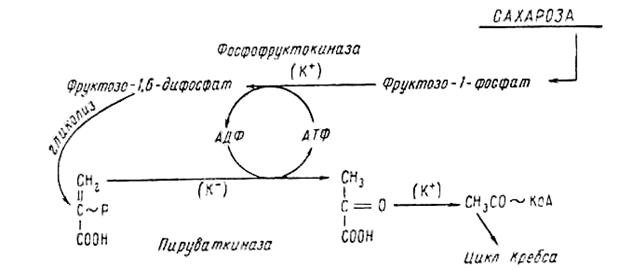
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
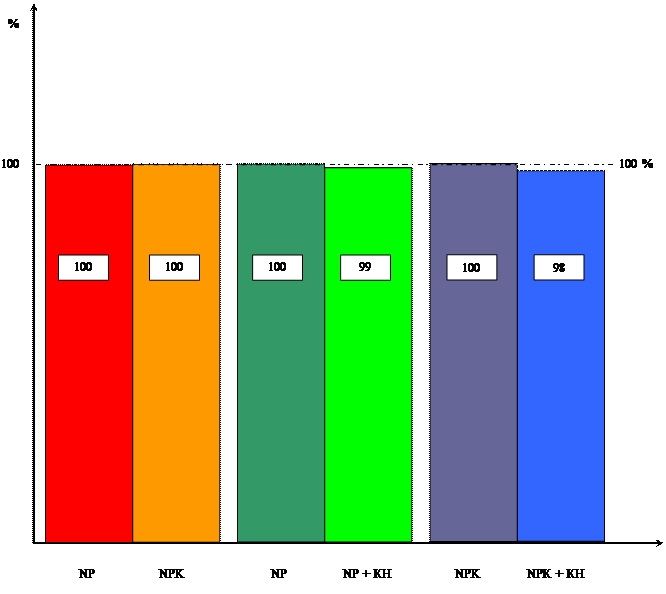
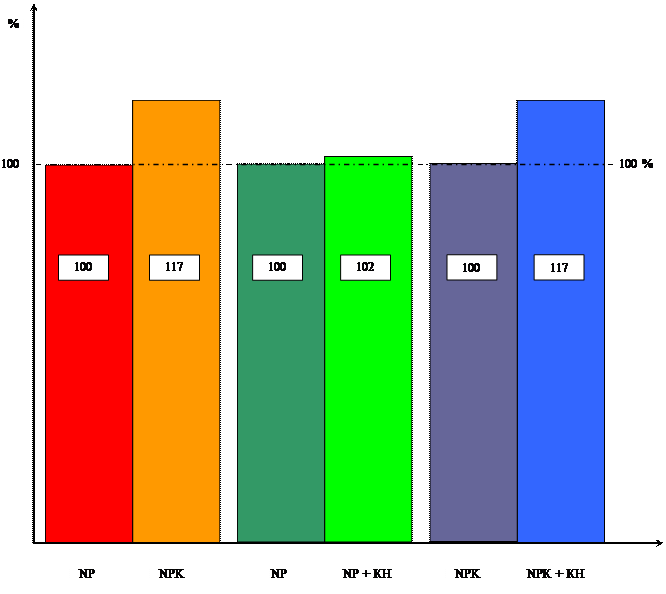
... системы, но вызвал увеличение ее массы, мы провели исследования по влиянию кинетина на накопление массы корневой и побеговой систем в зависимости от уровня минерального питания. Данные расположены в таблицах 3 и 4. Таблица 3 Влияние кинетина на массу побеговой системы 10-ти проростков пшеницы в зависимости от уровня калийного питания ( мг). Возраст, дн. NP NP + КН % NPK NPK + КН % ...
... или транспирация представляют совокупность химических и физических процессов. Чтобы понять механизм физиологического процесса, необходимо выделить его физические и химические компоненты. В связи с этим физиология растений все более нуждается в усовершенствованных методах биохимии. Биохимический подход оказался очень плодотворным в исследованиях таких сложных процессов, как фотосинтез и дыхание. ...
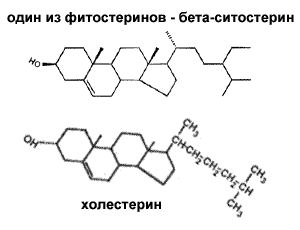
... токсичны для организма, а с другой — самостоятельный прием таких добавок может привести к нарушению баланса минеральных веществ в организме. Конечно есть и противопоказания, как в любом деле, касающемся здоровья человека. Для того чтобы их выявить, нужно поддерживать связь со своим врачом, а также подключить к составлению рациона (и заодно обсудить вопрос включения в него специальных “спортивных ...
0 комментариев