Навигация
СН3СООН ® СН3СОСН2СООН ® СН3СН2СН2СООН
274314
знаков
4
таблицы
0
изображений
2 СН3СООН ® СН3СОСН2СООН ® СН3СН2СН2СООН
Уксусная кислота используется для синтеза жирных кислот только в присутствии АДФ. Исходным соединением для биосинтеза жирных кислот ячвляется не сама уксусная кислота, а ацетилкофермент А, который дает активные ацетильные радикалы. Реакция происходит в два этапа по следующей схеме:
2 СН3СО»SКоА Ы СН3СОСН2СО»SКоА +НSКоА
СН3СОСН2СО»SКоА + Н2О Ы СН3СОСН2СООН + НSКоА
Образующаяся из ацетоуксусной кислоты масляная кислота, вступая в реакцию в ацетилКоА, образует капроновую кислоту. Однако, механизм биосинтеза высших жирных кислот у растений пока мало исследован.
Для осуществления синтеза жирных кислот необходимы также ионы марганца и НАДФ. Н2. В процессе синтеза жирных кислот участвует и углекислый газ, который вступает в реакцию с ацетилкоферментом А, образуя малонилкофермент А, являющийся важнейшим промежуточным продуктом при ферментативном синтезе жирных кислот.
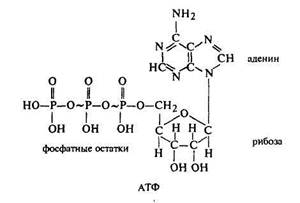
В процессе присоединения углекислого газа к ацетилкоферментуА важную каталитическую роль играет витамин биотин, а источником энергии для этого процесса является АТФ. Высшие ненасыщенные жирные кислоты образуются из насыщенных жирных кислот в процессе дегидрирования.
Процесс расщепления жира в растении происходит особенно энергично при прорастании семян. Наиболее ярко этот процесс выражен у масличных культур. Он начинается с гидролитического распада жиров, происходящего под действием фермента липазы и сопровождается накоплением глицерина и свободных жирных кислот, которые используются для различных синтезов в развивающемся проростке. При этом главным продуктом распада жиров является сахар. Из ненасыщенных жирных кислот сахара образуются быстрее.
Важную роль в метаболизме липидов играют такие ферменты, как сатураза (катализирует гидрирование ненасыщенных кислот в насыщенные) и липоксигеназа (катализирует окисление ненасыщенных кислот).
Важнейшим этапом диссимиляции жирных кислот является b-окисление, открытое Федором Кноопом. Процесс изучен в теме "Дыхание растений" в вопросе о глиоксилатном цикле. В этом процессе принимает участие кофермент А, происходит он, как правило, в митохондриях.
Наряду с b-окислением жирные кислоты подвергаются и a-окислению. При этом процесс окисления начинается с декарбоксилирования жирной кислоты под действием пероксидазы и при участии перекиси водорода, в результате образуется альдегид, содержащий уже на 1 атом углерода меньше, чем исходная жирная кислота, который под действием альдегиддегидрогеназы превращается в новую, более простую, жирную кислоту.
Связи между тремя основными группами органических веществ.
Поскольку все три основные группы органических веществ тесно связаны в метаболизме, можно выделить два основных ключевых момента в их взаимопревращении. Это прежде всего образование пировиноградной кислоты и уксусной кислоты. Именно эти два вещества являются теми краеугольными камнями, на которых основываются круговороты углеводов, жиров и белков.
От пировиноградной кислоты отходят пути образования глюкозы, а, следовательно и глюкозо-1-фосфата, как основы образования углеводов, и образование органических кислот (кетокислот), которые начинают путь синтеза аминокислот.
Уксусная кислота, образовываясь в русле синтеза органических кислот от пировиноградной кислоты, является началом пути образования жиров, а в русле расщепления жирных кислот в результате b-окисления, является связкой между метаболизмом жиров и углеводов.
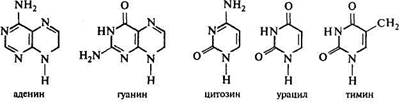
Образование нуклеиновых кислот, различных вторичных органических соединений основывается на веществах, синтезирующихся на промежуточных этапах синтеза этих трех групп веществ.
Передвижение органических веществ в растении.
В растении лист является основным органом биосинтеза. Продукты фотосинтеза запасаются в виде крахмала в хлоропластах и лейкопластах, перераспределение углеводов происходит при переходе крахмала в растворимые простые сахара.
В растении ксилема служит для перемещения воды и минеральных веществ из почвы в надземную часть, а флоэма служит для доставки сахарозы из листьев в другие органы растения.
По флоэме отток веществ наблюдается от донора (органа-синтезатора) вверх и вниз - к любому органу-акцептору, где эти вещества запасаются или потребляются. Органы, акцептирующие вещества, относятся, как правило, к запасающим органам (корнеплоды, корневища, клубни, луковицы).
По ксилеме же вещества движутся только снизу вверх.
Все потребляющие органы обеспечиваются, как правило, ближайшим к ним донором. Верхние фотосинтезирующие литься снабжают растущие почки и самые молодые листья. Нижние листья обеспечивают корни. Плоды обеспечиваются из ближайших к ним листьев.
Транспорт по флоэме может происходить одновременно в двух направлениях. Эта "двухнаправленность" является результатом одностороннего тока в отдельных, но смежных ситовидных трубках, соединенных с различными донорами и акцепторами.
Ситовидные трубки - это тонкостенные удлиненные клетки, соединенные своими концами и образующие непрерывную трубку. В местах соприкосновения клеточные стенки пронизаны ситовидными порами и называются поэтому ситовидными пластинками. В отличие от ксилемных клеток ситовидные флоэмные клетки - живые, хотя и непохожи на обычные живые клетки. Они не имеют ядра, но содержат некоторые другие органеллы и плазмалемму, которая играет важную роль в удержании сахаров в ситовидных трубках. Доказательством может служить способность флоэмных клеток к плазмолизу. Ситовидные трубки имеют короткий период жизни и постоянно заменяются новыми, образующимися при делении камбия.
Перемещение веществ по флоэме происходит с большой скоростью: до 100 см/час. Транспорт по флоэме осуществляется путем перетекания растворов. Высокое гидростатическое давление, обусловленное движением воды в богатые сахаром зоны с высоким отрицательным водным потенциалом, вызывает перетекание растворов в зоны с более низким давлением. Удаление сахара из них гарантирует постоянное наличие градиента и, следовательно, перетекание раствора. Загрузка растворенных веществ включает совместный транспорт (котранспорт) сахарозы и ионов водорода с участием специфической пермеазы. Этот процесс обусловлен градиентом кислотности и электрохимическим градиентом. Поглощенные ионы водорода выделяются впоследствии с помощью протонного транспортера, использующего энергию АТФ.
Кроме сахарозы во флоэмном потоке транспортируются аминокислоты и амиды (аспарагин, глютамин), при старении добавляются также органические и минеральные вещества из отмирающих органов.
В направленном транспорте ассимилятов в растении участвуют в основном три системы:
выталкивающая или нагнетающая (лист),
проводящая (флоэма),
аттрагирующая или притягивающая (меристематические и запасающие ткани).
Таким образом передвижение веществ в растении включает сложный комплекс процессов передвижения пасоки по ксилеме и флоэме, который регулируется растением и зависит как от внешних факторов, так и от фазы развития растения.
Тема: Рост и развитие растений.
Дополнительная литература:
М.Х. Чайлахян. Гиббереллины растений. Изд-во АН СССР, 1961, 63 с.
Ж. Бернье, Ж. - М. Кине, Р. Сакс. Физиология цветения. т.1-2, М.: Агропромиздат, 1985
В.В. Полевой, Т.С. Саламатова. Физиология роста и развития растений. Изд-во Ленинградского университета, Л., 1991, 239 с.506 с.
Вопросы к теме:
Характеристика факторов, определяющих закономерности роста и развития растений.
Рост растений (закономерности, типы).
Виды движения у растений.
Развитие растений (типы онтогенеза, этапы онтогенеза, особенности периода эвокации, особенности фазы покоя).
Теория старения и омоложения растений Кренке.
Особенности созревания продуктивных частей растений.
Использование регуляторов роста в практике сельского хозяйства.
Характеристика факторов, определяющих закономерности роста и развития растений.
Все ранее изученные процессы в совокупности определяют прежде всего осуществление основной функции растительного организма - роста, образования потомства, сохранения вида. Эта функция осуществляется через процессы роста и развития.
Жизненный цикл любого эукариотного организма, т.е. его развитие от оплодотворенной яйцеклетки до полного формирования, старения и гибели в результате естественной смерти, называется онтогенезом.
Рост - это процесс необратимого новообразования структурных элементов, сопровождающийся увеличением массы и размеров организма, т.е. количественное изменение.
Развитие - это качественное изменение компонентов организма, при котором имеющиеся формы или функции превращаются в другие.
На оба процесса оказывают влияние различные факторы:
внешние абиотические факторы окружающей среды, например солнечный свет,
внутренние факторы самого организма (гормоны, генетические признаки).
Благодаря генетической тотипотентности организма, определяемой генотипом, происходит строго последовательное образование того или иного типа тканей в соответствии с этапом развития организма. Образование определенных гормонов, ферментов, типов тканей в определенной фазе развития растения определяется обычно первичной активацией соответствующих генов и называется дифференциальной активацией гена (ДАГ).
Вторичная активация генов, а также их репрессия могут происходить и под воздействием некоторых внешних факторов.
Одним из важнейших внутриклеточных регуляторов активации генов и развития того или иного процесса, связанного с ростовыми процессами или переходом растения в следующую фазу развития являются фитогормоны.
Изученные фитогормоны делят на две большие группы:
стимуляторы роста
ингибиторы роста.
В свою очередь стимуляторы роста делят на три класса:
ауксины,
гиббереллины,
цитокинины.
К ауксинам относятся вещества индольной природы, типичным представителем является индолил-3-уксусная кислота (ИУК). Образуются они в меристематических клетках и передвигаются как базипетально, так и акропетально. Ауксины ускоряют митотическую активность как апикальных меристем, так и камбия, задерживают опадание листьев и завязей, активируют корнеобразование.
К гиббереллинам относятся вещества сложной природы - производные гибберелловой кислоты. Выделены из аскомицетных грибов (род Gibberella fujikuroi), имеющих ярко выраженную конидиальную стадию (род фузариум). Именно в конидиальной стадии этот гриб вызывает у риса болезнь "дурных побегов", характеризующуюся стремительным ростом побегов, их вытягиванием, истончением, и, как следствие, гибелью. Гиббереллины также транспортируются в растении акропетально и базипетально как по ксилеме, так и по флоэме. Гиббереллины ускоряют фазу растяжения клеток, регулируют процессы цветения и плодоношения, индуцируют новообразование пигментов.
К цитокининам относятся производные пурина, типичным представителем которых является кинетин. Эта группа гормонов не обладает столь ярко выраженным действием, как предыдущие, однако цитокинины воздействуют на многие звенья метаболизма, усиливают синтез ДНК, РНК, белков.
Ингибиторы роста представлены двумя веществами:
абсцизовая кислота,
этилен.
Абсцизовая кислота - это гормон стресса, ее количество сильно увеличивается при недостатке воды (закрывание устьиц) и питательных веществ. АБК подавляет биосинтез нуклеиновых кислот и белков.
Этилен - это газообразный фитогормон, который тормозит рост и ускоряет созревание плодов. Этот гормон выделяется созревающими органами растений и воздействует как на другие органы этого же растения, так и на растения, находящиеся рядом. Этилен ускоряет опадение листьев, цветков, плодов за счет освобождения целлюлазы у черешков, что ускоряет образование отделительного слоя. Этилен образуется при распаде этрела, что сильно облегчает его практическое применение в сельском хозяйстве.
Рост растений (закономерности и типы).
Под термином рост у растений подразумевается несколько процессов:
рост клетки,
рост ткани,
рост растительного организма в целом.
Рост клетки характеризуется наличием следующих фаз:
Эмбриональная фаза (вакуоли нет, остальные органоиды в небольшом количестве).
Фаза растяжения (появление вакуоли, укрепление клеточной стенки, увеличение размера клетки).
Фаза дифференциации (появление в клетке специфических для данной ткани органоидов).
Рост ткани в зависимости от ее специфичности может проходить по какому-либо из типов:
Апикальному (побег, корень).
Базальному (лист).
Интеркалярному (стебель у злаков).
Рост растительного организма в целом характеризуется наличием следующих фаз:
Лаг-фаза или индукционный рост (прорастание семени).
Лог-фаза или фаза логарифмического роста (формирование вегетативной массы растения).
Фаза замедленного роста (в период плодоношения, когда образование новых вегетативных частей растения ограниченно).
Фаза стационарного состояния (совпадает, как правило, со старением и отмиранием растения).
Скорость роста и относительный рост или прирост у растений определяют, измеряя параметры растений в определенном временном режиме.
Для определения прироста пользуются разнообразными методами, в частности:
с помощью линейки,
с помощью горизонтального микроскопа,
с помощью меток,
с помощью ауксанографа,
с помощью масштабного фотографирования.
В среднем скорость роста у растений составляет 0,005 мм/мин., однако есть быстрорастущие растения и органы: тычинки злаков растут со скоростью 2 мм/мин., бамбук - 1 мм/мин.
По результатам современных исследований (В.С. Шевелуха) предложена следующая классификация типов роста:
синусоидальный тип (кривая суточного хода скорости линейного роста имеет вид синусоиды с фазой максимума в дневные и минимума в ранние утренние часы) (характерна для злаков),
импульсный тип роста (кривая усиления скорости ростовых процессов и их торможения происходит скачкообразно под прямым или острым углом в течение десятков минут. Максимальная скорость роста наступает в 20-21 час и сохраняется всю ночь, днем рост заторможен) (характерна для корнеплодов и клубней),
двухволновой тип (в течение суток скорость роста имеет две волны, дважны достигая максимума и минимума),
выравненный тип роста (кривая роста имеет плавный характер).
Виды движения у растений.
Несмотря на то, что растения, как правило, стационарно закрепляются в окружающем пространстве, они способны к ряду видов движения.
Основные виды движения у растений:
таксисы.
нутации,
настии,
тропизмы,
Таксисы характерны только для низших водных неприкрепленных растений,
для высших растений характерны первые три вида.
Нутации совершают растущие апикальные побеги, вращаясь вокруг своей оси, причем надземные побеги совершают их только под влиянием гормонов, а корни - как под влиянием гормонов, так и с помощью особых клеток (статоцитов (с органеллами статолитами), которые способны использовать естественные силы гравитации при осуществлении этого процесса.
Настии растение совершает под влиянием равномерно действующего абиотического фактора (света, воды и т.д.).
Тропизмы растение совершает под влиянием неравномерно действующего абиотического фактора (света, воды, гравитации и т.д.).
Развитие растений (типы онтогенеза, этапы онтогенеза, особенности периода эвокации, особенности фазы покоя).
Развитие растений или онтогенез характеризуется тем, что на переход растения из одной фазы онтогенеза в другую действуют очень большое количество факторов, причем часто необходимо их совокупное действие.
Различают следующие типы онтогенеза растений:
По продолжительности жизни:
однолетние,
двулетние,
многолетние;
По количеству плодоношений:
монокарпические,
поликарпические.
Любое растение проходит в процессе онтогенеза следующие этапы развития:
эмбриональную фазу (от оплодотворения семяпочки до формирования семени),
ювенильную фазу (от прорастания семени до появления всхода на поверхности почвы),
фазу формирования надземных вегетативных органов,
фазу цветения и плодообразования,
фазу созревания,
фазу отмирания.
Наиболее насыщенной является ювенильная фаза развития, которая делится на такие периоды, как:
набухание,
проклевывание,
гетеротрофный рост проростка в темноте,
переход к автотрофному типу питания.
Практически каждое онтогенетическое изменение происходит под воздействием внутренних и внешних факторов. При этом из внешних фактором наиболее важное значение имеет солнечный свет. Переход к автотрофному способу питания, переход к фазе бутонизации и цветения, переход к состоянию покоя у многолетних растений непосредственно связаны именно с воздействием продолжительности солнечного освещения и поэтому называются фотоморфогенезом. Свет является сигналом не тольлко к смене фазы развития, но и непосредственно влияет на рост, транспирацию и другие физиологические процессы в растении. Непосредственное воздействие света выражается в способности клеток образовывать соответствующие гормоны, в частности абсцизовую кислоту, что позволяет растению замедлять скорость роста при переходе к автотрофному питанию. Опосредованное воздействие света в виде длительности светового дня определяет переход к следующей фазе развития, в частности к цветению.
Восприятие растением воздействия солнечного света происходит благодаря наличию специальных фоторецепторов и гормонов.
Непосредственное воздействие света воспринимается растением с помощью фоторецептора "криптохром", и пигмента "фитохром". Особенно важен фитохром, который способен воспринимать различные составляющие спектра солнечного света и, в зависимости от поглощенной длины волны, превращается либо в форму Фк, поглощающий красный свет с длиной волны 600 нм, либо в форму Фдк, поглощающий дальний красный свет с длиной волны 730 нм. При обычных условиях этот пигмент находится в обеих формах в равных пропорциях, однако, при смене условий, например на затененные, происходит образование большего количества пигмента Фк, и это определяет вытягивание и этиолирование тканей побега. На основе действия этих фоторецепторов и пигментов растение проходит суточные изменения в определенном ритме, который называется циркадным, или биологическими часами растения.
Световой фактор вызывает также синтез определенных гормонов, которые определяют переход растения в фазу цветения или в фазу эвокации, т.е. переход от вегетативного состояния к генеративному развитию. Основным гормоном, действующим на этом этапе онтогенеза, является гормон "флориген", состоящий из двух групп гормонов:
гиббереллинов, вызывающих образование и рост цветоносов,
антезинов, вызывающих формирование цветков.
Понимание этого момента весьма важно на практике, особенно в плодоводстве, где использование подвоя и привоя в определенных фазах онтогенеза будет влиять на скорость вступления в плодоношение привитого растения. Поток гормонов, в том числе и флоригена, идет от привоя к подвою, поэтому важно использование подвоя с растения, находящегося в определенной фазе развития. Флоральный морфогенез контролируется сложной системой многих факторов, каждый из которых в необходимой концентрации и в нужное время запускает свою цепь процессов, ведущих к закладке цветков.
Вторым важным фактором, играющим определенную роль в формировании флорального морфогенеза является температурный фактор. Он особенно важен для озимых и двулетних культур, поскольку именно пониженные температуры вызывают у этих культур те биохимические преобразования, которые определяют синтез флоригена и других сопутствующих гормонов, определяющих инициацию цветения.
Именно на действии пониженных температур основан прием яровизации, который используется в различных опытных исследованиях, когда необходимо ускорить смену поколений у озимых культур. К таким же результатам приводит и обработка растений гиббереллинами, благодаря которой можно ускорить цветение двулетних растений.
По отношению к фотопериоду растения делят на три группы:
растения короткого дня (цветение при длине дня меньше 12 часов) (хризантема, георгин, топинамбур, просо, сорго, табак),
растения длинного дня (цветение при длине дня больше 12 часов) (астра, клевер, лен, лук, морковь, свекла, шпинат),
нейтральные растения (цветение не зависит от длины дня) (подсолнечник, гречиха, бобы, рапс, томат).
В онтогенезе растений обязательно имеется фаза ослабления жизнедеятельности, которая носит название состояния покоя. У однолетних растений это состояние наступает только один раз - при формировании семени, у многолетних растений - много раз при переходе к существованию в неблагоприятные условия среды (зима, засуха). Покой - это такое состояние растения, которое характеризуется отсутствием ростовых явлений, крайней степенью угнетенности дыхания и снижением интенсивности превращения веществ.
Различают летний и зимний покой у многолетников, глубокий и вынужденный покой у всех растений. Вынужденный покой возможен только при участии человека, который может обеспечить особые условия хранения покоящихся органов в специальных хранилищах с помощью специальных методов. Очень важным моментом перехода в состояние покоя является этап послеуборочного дозревания, что позволяет предотвратить преждевременное прорастание семян, сконцентрировать максимальное количество запасных веществ.
Теория старения и омоложения растений Кренке.
В процессе онтогенеза растение подвергается определенным изменениям, которые связаны с явлением возрастной изменчивости. Теорию, объясняющую закономерности этой изменчивости предложил в 40 годы прошлого столетия, Н.П. Кренке. Основные постулаты этой теории:
Каждый организм, начиная от возникновения, непрерывно стареет до своей естественной смерти.
В первой половине жизни старение прерывается периодически омоложением, т.е. образованием новых побегов, листьев и т.п., что замедляет темп старения.
Растениям присущ физиологический возраст, который определяет истинный возраст органа растения: листья однолетнего и десятилетнего деревьев неравноценны, неравноценны и листья на одном дереве, но на побегах разного порядка. Различают понятие "возраст" (календарный возраст) и "возрастность" (физиологический возраст. Возрастность определяется возрастом органа и материнского растения. В пределах плодового дерева листья на побегах высших порядков ветвления физиологически более старые, чем листья того же возраста на побегах низших порядков ветвления. Поэтому по форме, анатомическому строению, физиологическим и биохимическим признакам верхние листья, несмотря на свой меньший возраст, обнаруживают признаки большего старения, срок их жизни часто короче, чем у средних листьев на том же побеге.
Цикличность онтогенетического развития заключается в том, что дочерние клетки при своем новообразовании являются временно омоложенными по отношению к материнским.
Скорость старения и нормальная средняя продолжительность жизни определяются начальным потенциалом жизнеспособности и обуславливаются генетическими особенностями вида.
Проблемой старения и омоложения плодово-ягодных культур занимался и П.Г. Шитт. В 60-ых годах прошлого столетия он впервые установил наличие возрастных качественных изменений у корней. И.В. Мичурин также указывал на тесную связь между органообразовательными процессами в организмах и возрастной изменчивостью.
Установленные Н.П. Кренке закономерности изменения морфологии листьев и побегов в связи с их возрастом позволили разработать рекомендации по ранней диагностике скороспелости растений в пределах вида, выявить коррелятивные связи между качеством клубней и корнеплодов и скороспелостью сорта. Установлено, что для скороспелых сортов характерно резкое изменение морфологических признаков листьев (быстрое пожелтение и отмирание листьев), а у позднеспелых сортов изменения происходят постепенно. Эта закономерность имеет важное значение в процессе селекции сортов на скороспелость и качество.
Морфологические признаки тесно связаны с генетически заложенной скороспелостью, что дает возможность использовать их в селекции плодовых культур, например:
у однолетних сеянцев скороспелых сортов яблони междоузлия короче, ветвление более сильное, листья расположены гуще, чем у сортов, позже вступающих в плодоношение,
у двулетних сеянцев яблони интенсивность зеленой окраски листьев при переходе от верхних ярусов к нижним у скороспелых форм изменяется более резко, чем у позднеспелых,
чем выше по стеблю плодового растения взят черенок или почка (при вегетативном размножении), тем раньше после укоренения или окулировки растение способно зацветать.
На основе теории Кренке были усовершенствованы приемы подрезки растений, технология выбора побегов и их частей требуемого качества при вегетативном размножении растений, обеспечивающая лучшую укореняемость черенков, технология достижения оптимального сочетания вегетативного и генеративного развития растений при черенковании и прививках.
Особенности созревания продуктивных частей растений.
Продуктивными частями растений называют как органы генеративного размножения (плоды, семена), так и органы вегетативного размножения (клубни, луковицы). Остальные продуктивные части (листья у зеленных культур, стеблеплоды, корнеплоды и др.) не несут функции размножения и поэтому закономерности из роста и развития не имеют столь важного значения.
Плод, содержащий семена, является органом размножения у подавляющего большинства сельскохозяйственных растений. Он выполняет две функции:
защита семян,
распространение семян.
Для осуществления этих функций у различных плодов имеются соответствующие приспособления (сухие и сочные плоды, крючочки, крылатки, привлекательный вкус и т.д.).
В развитии плода выделяют четыре фазы:
Формирование завязи до опыления,
Рост за счет деления клеток сразу после опыления и оплодотворения,
Рост за счет растяжения клеток,
Созревание.
Рост завязи стимулируется прорастающей пыльцой еще до образования зиготы, причем интенсивность этого роста прямо пропорциональна количеству прорастающей пыльцы. Даже чужеродная пыльца может способствовать росту завязи, что объясняется высоким содержанием ИУК в пыльце.
Обработка цветков экзогенным ауксином у многих растений с сочными плодами индуцирует разрастание завязи и образование партенокарпических, т.е. бессемянных плодов. Обработка гиббереллином также у многих растений вызывает завязывание плодов (виноград, яблоня, томат и т.п.). Для роста молодых плодов необходимо присутствие цитокинина, но экзогенные цитокинины не вызывают образования партенокарпических плодов.
В начале формирования завязи в цветке ее рост происходит в результате деления клеток, которое резко возрастает после опыления. Затем наступает более продолжительная фаза растяжения клеток. Характер роста находится в сильной зависимости от типа плода.
Регуляция роста плодов осуществляется фитогормонами. ИУК в завязь сначала поступает из столбика и от прорастающей пыльцы. Затем источником ИУК становится развивающаяся семяпочка. При этом играет определенную роль и гормон старения (этилен), который обеспечивает увядание цветка после опыления. Образующиеся семена поставляют ауксин в околоплодник, что активирует в нем ростовые процессы. При недостатке ауксина (малочисленности образующихся семян) происходит опадение плодов.
Содержание фитогормонов и их соотношение различны на разных фазах роста плодов и формирования семян.
Так, в зерновках пшеницы максимальное количество цитокининов наблюдается сразу после цветения при переходе к формированию эндосперма. Затем начинает возрастать содержание гиббереллинов, а позже ИУК, концентрация которых достигает максимальной величины в фазе молочной спелости. При переходе к восковой спелости уровень гиббереллинов и ауксинов быстро падает, но возрастает содержание АБК, что способствует отложению в эндосперме запасных веществ. Когда нарастание сухой массы зерновок прекращается и происходит обезвоживание семян (полная спелость), содержание АБК снижается. Уменьшение количества всех фитогормонов объясняется переходом их в связанное состояние. Такой порядок изменения соотношения фитогормонов у формирующихся зерновок пшеницы обуславливается последовательность развития зародыша и эндосперма. При созревании зерновки накапливаются углеводы и белки, происходят изменения в нуклеиновом обмене, пластические вещества активно перемещаются в зерновки из стеблей и листьев. Происходит раздревеснение стеблей (снижается содержание клетчатки, лигнина, которые преобразуются в крахмал). При созревании зерна белок становится более устойчивым к действию протеолитических ферментов, уменьшается количество моносахаридов и увеличивается количество крахмала.
Бобовые культуры накапливают значительно меньше крахмала и других углеводов, чем злаковые культуры.
При возделывании зерновых и зернобобовых культур часто применяется раздельный способ уборки, который позволяет лучше обеспечить переход пластических веществ из стеблей в семена после скашивания и дозаривания в валках. Обработка посевов в период восковой спелости раствором азотнокислого аммония ускоряет созревание этих культур на 5-7 дней.
При созревании семян масличных культур жиры не только накапливаются, но и меняются в качественном отношении. В недозрелых семенах содержится больше свободных и насыщенных жирных кислот, в зрелых - увеличивается содержание ненасыщенных жирных кислот.
В сочных плодах наибольшее содержание гиббереллинов и ауксина в перикарпе наблюдается в начале его развития. Затем уровень этих фитогормонов снижается и снова возрастает на последней фазе роста. Содержание цитокинина временно увеличивается в период наиболее интенсивного роста плода. Прекращение роста перикарпа совпадает с накоплением АБК в его тканях.
Период растяжения клеток у сочных плодов и особенно конец этого периода характеризуются не только интенсивным ростом, но и накоплением органических веществ. Происходит увеличение содержания углеводов и органических кислот, откладывается крахмал.
Созревание некоторых плодов хорошо коррелирует с увеличением скорости дыхания. Период повышенного выделения углекислого газа плодом называется климактерием, и в этот период плод претерпевает изменение из незрелого в созревший. Обработка этиленом стимулирует этот период и дозревание спелых плодов. Этилен повышает проницаемость мембран в клетках плода, что дает возможность ферментам, ранее отделенным от субстратов мембранами, вступить в реакцию с этими субстратами и начать их разрушение.
Ауксин также участвует в созревании плода, причем при созревании плодов и опадении листьев ауксин и этилен действуют как антагонисты. Какой гормон доминирует при этом, зависит от возраста ткани.
У ряда культурпреобладающим способом размножения стал именно способ размножения с помощью органов вегетативного размножения (например, картофель). Поэтому формирование этих органов, как выполняющих и репродуктивную функцию, и, в то же время, служащих в качестве источника питания для человека, требует отдельного рассмотрения.
Процесс клубнеобразования в физиологическом плане лучше всего изучен у картофеля. При длинном дне и высокой температуре (свыше 29 градусов) могут превращаться в вертикальные облиственные побеги, а при нормальной (более низкой) температуре на конце столона формируется клубень. Клубнеобразование всегда связано с торможением роста как надземных побегов, так и столонов. Короткий день способствует поступлению в клубни пластических веществ.
Формирование клубня включает в себя три этапа;
подготовительный - появление и рост столонов,
закладку и рост самого клубня,
дозревание и покой клубня.
Образованию столонов из пазушных почек благоприятствует их затемнение (именно поэтому в технологии возделывания картофеля обязателен прием окучивания). ИУК вместе с гиббереллинами, поступающие в достаточном количестве из надземных частей, переключают генетическую программу развития пазушной почки с развития вертикального облиственного побега на формирование столона. Гиббереллин необходим и для удлинения междоузлий столона.
Закладка клубней на дистальных концах столонов связана с резким торможением их роста в длину. По-видимому, это подавление вызывается повышением концентрации АБК, которая в большом количестве образуется в листьях на коротком дне. В условиях короткого дня снижаются синтез и поступление ИУК и гиббереллинов. Одновременно увеличивается отношение цитокининов в ауксинам.
Покой клубней связан с резким замедлением дыхания, распада и синтеза биополимеров, остановкой ростовых процессов. У клубней картофеля в состоянии глубокого покоя находятся только меристематические ткани, прежде всего глазки. Запасающая ткань способна быстро активизироваться в ответ на повреждения (образуется раневая перидермы при механическом повреждении).
Состояние глубокого покоя глазков обусловлено высоким содержанием АБК, кофейной кислоты и скополетина.
Выход глазков из состояния глубокого покоя связан с падением содержания АБК (в 10-100 раз) и возрастанием концентрации свободных гиббереллинов. Обработка стимуляторами на основе гиббереллиновой кислоты прекращает состояние покоя у клубней и позволяет проводить летние посадки картофеля на юге.
У луковиц в период покоя ростовые процессы не прекращаются, хотя они идут очень медленно. Состояние покоя поддерживается высокой концентрацией АБК. Перед прорастанием уровень АБК снижается, а содержание цитокининов, гиббереллина и ауксинов возрастает.
Таким же закономерностям в смене работы разных фитогормонов подчиняются процессы образования корневищ и столонов, а также способность растений укореняться с помощью отводков и черенков.
Использование регуляторов роста в практике сельского хозяйства.
Регуляторы роста достаточно широко используются в практике сельского хозяйства в следующих направлениях:
На стадии посева, посадки,
На стадии управления цветением, завязыванием, формированием урожая,
На стадии уборки,
На стадии покоя.
На стадии посева, посадки используют:
Ауксины:
для укоренения трудноукореняемых черенков, например винограда,
для лучшей приживаемости прививок,
для лучшего прорастания семян
На стадии управления цветением, завязыванием, формированием урожая используют:
Ауксины:
для стимулирования начала цветения,
для увеличения количества завязываемых плодов,
для стимулирования женского цветения у раздельнополых видов.
Гиббереллины:
для увеличения размера плодов,
для улучшения качества хозяйственно-ценных органов (способствуют увеличению сахаров в плодах, стеблях, стеблеплодах, корнеплодах и т.д.),
для стимулирования мужского цветения у раздельнополых видов.
Этилен и абсцизовая кислота также стимулируют женское цветение у раздельнополых видов.
На стадии уборки используют:
Этилен и абсцизовую кислоту и ряд других ингибиторов роста (например: хлорат магния, гидрел, этрел):
для ускорения созревания, повышения дружности отдачи урожая,
для дефолиации,
для десикации (предуборочное высушивание стеблей и листьев),
для сеникации (ускорение созревания на 5-7 дней в районах с коротким теплым периодом)
На стадии покоя:
Для продления состояния покоя используют этилен и абсцизовую кислоту для обработки продовольственного картофеля, корнеплодов, плодов (либо опрыскивают 0,5% раствором гидрела, либо регулируют состав атмосферы в хранилище),
Для нарушения состояния покоя используют:
эфиризацию: для прорастания побегов, корневищ - обработку серным эфиром,
теплые ванны: для выгонки сирени к Новому году (опускают побеги куста в теплую (30-35оС) воду на 9-12 часов),
гиббереллины для получения второго урожая картофеля от свежеубранных клубней (из замачивают на 30 минут в смеси 0,0005% гиббереллина и 2% тиомочевины).
Тема: Устойчивость растений к факторам окружающей среды.
Дополнительная литература:
А.А. Жученко. Экологическая генетика культурных растений. Кишинев, "Штиинца", 1980,
С.А. Блинкин, Т.В. Рудницкая. Фитонциды вокруг нас. Изд-во "Знание", М, 1981.
Адаптация растений к экстремальным условиям увлажнения (обзор), Кишинев, "Штиинца", 1984,
Ф.Д. Сказкин. Критический период у растений по отношению к недостатку воды в почве. Л., Наука, 1971,
М.Д. Кушниренко. Физиология засухоустойчивости плодовых растений. Кишинев, Штиинца, 1975,Экспресс-методы диагностики жаро-, засухоустойчивости и сроков полива растений. Кишинев, Штиинца, 1986,
И.А. Буркин. Защита овощных культур от заморозков. М., Россельхозиздат, 1973,
Холодостойковть растений. Под ред. Г.А. Самыгина, М., "Колос", 1983,
Г.В. Удовенко. Состояние и пути решения проблемы солеустойчивости растений. М., 1978.
Вопросы к теме:
Основные способы приспособления растений к факторам окружающей среды.
Приспособление растений к температурному фактору и недостатку влаги.
Причины возникновения полегания и меры борьбы с ним.
Приспособление растений к уровню кислотности почвы,
Приспособление растений к повышенному уровню засоления.
Приспособление растений к содержанию загрязняющих атмосферу газов.
Приспособление растений к биотическим факторам - болезням, вредителям.
Основные способы приспособления растений к факторам окружающей среды.
Факторы окружающей среды, действующие на растения, делятся на абиотические и биотические. По отношению к этим двум группам факторов у растений выработались в процессе эволюции своеобразные методы защиты или приспособления к их переживанию.
Так по отношению к абиотическим факторам различают три главных способа приспособления растений:
механизмы, позволяющие избежать неблагоприятное воздействие (переход в состояние покоя: образование почек, семян, вегетативных органов),
специальные структурные приспособления (различные видоизменения листьев, стеблей цветков и т.д.),
физиологические механизмы (С4-путь фотосинтеза, САМ-фотосинтез, увеличение вязкости цитоплазмы).
По отношению к биотическим факторам у растений также различают несколько способов приспособления (естественной защиты):
приспособления морфологического характера,
механизм неспецифического иммунитета, т.е. приспособления биохимического характера (фитонциды, фитоалексины, алкалоиды),
механизм специфического иммунитет (выработка специализированных антител против возбудителей болезней).
Приспособление растений к температурному фактору.
Существенное действие на растения оказывают как низкие температуры, так и высокие температуры.
По отношению к низким температурам различают:
холодостойкость, т.е. способность растения переносить низкие положительные температуры, при этом у растений не происходят изменения в ферментативном аппарате, поскольку сами ферменты, по-видимому, имеют структурные особенности, позволяющие сохранить пространственную структуру и биологическую активность в низкотемпературных условиях,
морозостойкость, т.е. способность растений переносить охлаждение ниже ОоС,
зимостойкость, т.е. комплексная устойчивость растений против неблагоприятных факторов зимы (морозов, чередующихся с оттепелями, ледяной корки, снеговалов и т.п.).
При переживании растениями зимних условий особое значение имеют типичные повреждения растений и способы профилактики этих повреждений:
выпирание,
выпревание,
вымокание,
витрификация.
Выпирание - это гибель озимых культур, наступающая вследствие того, что в почве образуется ледяная прослойка, поднимающая верхний слой почвы вместе с растениями, что приводит к обрыву корневой системы. Ледяная прослойка образуется в том случае, когда талая вода, успевшая просочиться в почву, замерзает с наступлением морозов.
Выпревание - это гибель растений, находящихся под глубоким покровом снега в условиях мягкой зимы. Причина выпревания - расходование на дыхание запасенных с осени веществ без их фотосинтетического пополнения. Устойчивость к выпреванию определяется большим накоплением углеводов и низким уровнем дыхания в зимний период.
Вымокание - это гибель растений, происходящая преимущественно в весенний период или в период продолжительных оттепелей, когда на поверхности почвы скопляется талая вода, не впитавшаяся в замерзшую почву. Причиной гибели является недостаток кислорода. При замерзании этой талой воды образуется ледяная корка, которая может сдавливать и разрывать вмерзшие в нее растения озимых культур.
Витрификация - это переход свободной воды в клетках растения в стеклообразное состояние при резком охлаждении растений ниже 20 градусов мороза. Застывшая растительная ткань в виде аморфной стекловидной массы долго сохраняет свою жизнеспособность. Условия разморожения витрифицированных растений определяют возможность восстановления жизнеспособности:
при медленном отогревании ткани восстанавливаются,
при быстром отогревании происходит гибель клеток из-за того, что вода в большом количестве, поступающая в протопласт, не успевает перейти в связанное с белками состояние и повреждает структуры ядра и цитоплазмы.
Растения или органы растений, находящиеся в состоянии глубокого покоя, переносят очень низкие температуры, например в экспериментальных условиях удалось сохранить жизнеспособность черенков смородины после замораживания их до -253оС.
И.И. Туманов в 60-е годы прошлого века обосновал теорию закаливания растений, согласно которой при закаливании в тканях растений физиологические процессы идут с клетках в следующем порядке:
на первой фазе закаливания происходит накапливание сахаров, снижается осмотическое давление под влиянием низких положительных температур, прекращается рост растения,
на второй фазе закаливания при отрицательных температурах от 0 до -1оС, т.е. при температурах, еще не вызывающих необратимых повреждений клетки, наблюдается частичная потеря воды клетками, возрастает количество коллоидно-связанной воды.
Закаливанию растений способствует накопление ингибиторов роста, а ослабляет процесс закаливания увеличение концентрации гиббереллинов.
На проявление морозоустойчивости растений оказывает определенное влияние фотопериод в данных климатических условиях, например, длинный день способствует накоплению стимуляторов роста, а короткий день - ингибиторов роста.
Устойчивость к заморозкам у сельскохозяйственных культур различна.
| Сельскохозяйственные культуры | температурный предел, который они выдерживают в фазе всходов |
| горох, капуста, овес, пшеница, ячмень | до -10оС |
| бобы, морковь, подсолнечник, свекла | до -8оС |
| картофель, кукуруза, махорка | до -3оС |
| арахис, бахчевые, гречиха, огурец, табак, томат, фасоль | до +1оС |
В практике сельского хозяйства применяют следующие приемы повышения холодостойкости растений:
закалка набухших семян в течение 5-10 дней попеременно температурами выше и ниже 0оС,
намачивание семян теплолюбивых культур (кукурузы) в растворе алюмокалиевых квасцов,
замачивание семян в растворах микроэлементов, азотнокислого аммония,
закалка рассады в теплицах и парниках низкими положительными температурами (открывание рам, поднятие "фартуков" в пленочных теплицах днем).
Повышение морозостойкости при выращивании южных плодовых культур в северных районах достигают путем искусственного сокращения длины дня, при этом укорачивается период вегетации и создаются условия для вызревания древесины, полного ухода в состояние покоя почек, а, следовательно, обеспечивается успешная перезимовка.
Большое значение имеет холодо - и морозостойкость у озимых культур. Узел кущения у озимых культур - это единственный орган, способный к образованию новых корней и надземных побегов, поэтому особое значение имеет его глубина залегания. У зимостойких сортов эта величина в 1,5-2 раза больше, чем у не зимостойких сортов. С увеличением глубины заделки семян при посеве возрастает и глубина залегания узла кущения.
Таким образом можно управлять процессом перезимовки, регулируя сроки, способы посева, глубину посева, нормы высева, активно применяя весеннее боронование, стимулирующее способность к регенерации узла кущения злаков, подбирая сорта, наилучшим образом приспособленные к данным климатическим условиям.
По отношению к высоким температурам различают:
жаростойкость - способность растений переносить перегрев от 40 до 65оС. При повреждении растений высокими температурами без изменения уровня влажности в тканях происходит разрушение белково-липидного комплекса мембран, клетки теряют осмотические свойства, происходит обезвоживание тканей, разрушение коллоидных структур цитоплазмы.
засухоустойчивость - способность растений переносить атмосферную и почвенную засуху.
Засухи делятся на следующие виды в зависимости от времени наступления:
весенняя засуха характеризуется сравнительно низкими температурами, низкой относительной влажностью воздуха и сильными сухими ветрами,
летняя засуха характеризуется высокими температурами, низкой влажностью воздуха и повышенной испаряемостью,
осенняя засуха характеризуется высокими температурами, низкой влажностью воздуха и сильным иссушением почвы.
В зависимости от особенностей протекания засухи делятся на два типа:
1. Почвенная засуха - характеризуется постепенным иссушением почвы и растения в некоторой степени способны временно приспособиться к ней. При этом виде засухи завядают сначала нижние листья. Но при достаточно длительном периоде продолжительности почвенной засухи в конце лета и осенью, она гораздо опаснее кратковременной атмосферной засухи.
Атмосферная засуха - характеризуется резким уменьшением относительной влажности воздуха (до 10-20%), вследствие чего корневая система не успевает подавать воду в листья при сильно увеличившейся транспирации. Происходит быстрое обезвоживание верхних листьев, репродуктивных органов. Для атмосферной засухи характерны такие явления, как "запал" и "захват". Запал - это повреждение листьев и других надземных органов под действием перегрева и высокой сухости воздуха. При сильной степени запала листья высыхают, оставаясь зелеными, при обычном запале хлорофилл распадается и листья буреют. Захват - это повреждение хлебов суховеями в стадии молочной или молочно-восковой спелости (конец июня - начало июля). Вследствие перегрева и резкого водного дефицита зерновки пересыхают, белки сворачиваются, нарушается отток пластических веществ из листьев и соломы. зерно щуплое с низкой всхожестью. Поэтому в зоне суховеев используют скороспелые сорта и проводят агролесомелиоративные мероприятия, снижающие интенсивность суховеев.
Ко всем видам воздействия температурным фактором растения приспосабливаются используя все виды приспособлений (морфологические - для снижения транспирации, физиологические - для изменения вязкости цитоплазмы, переход в состояние покоя).
Устойчивость растений к засухе или к повышенным температурам определяется различными методами как лабораторными (в специальных опытах в использованием отдельных тканей и органов), так и лабораторно-полевыми (в специальных опытах в вегетационных сосудах с контролируемым режимом полива). В полевых условиях засухо - и жаростойкость растений и необходимость полива определяют с помощью различных приборов:
"тургомера" (принцип действия - измерение толщины листьев при воздействии высокой температуры),
ЭСТЛП-1а (прибор для определения электрического сопротивления тканей листьев) (принцип действия - измерение электрического сопротивления тканей листьев, которое имеет тесную отрицательную сопряженность с содержанием воды в листьях и положительную сопряженность с водоудерживающей способностью листьев).
По отношению к температурному фактору и водному режиму все растения подразделяются на следующие экологические типы:
1. Ксерофиты (приспособлены к атмосферной засухе) и делятся на подтипы:
суккуленты (стойки к перегреву, содержат большое количество воды и медленно ее расходуют - кактусы, алоэ, очиток),
эвксерофиты (хорошо переносят засуху за счет морфологических и физиологических приспособлений - верблюжья колючка, полынь),
гемиксерофиты (полуксерофиты) (переносят обезвоживание и перегрев за счет глубокой корневой системы - шалфей),
стипаксерофиты (выносят перегрев, но почвенную засуху переносят плохо - ковыль и другие степные злаки),
пойкилоксерофиты (уходят от неблагоприятных условий - степные тюльпаны, лишайники),
Похожие работы

... , в которых выращивают растения и проводят опыты в условиях определенного состава воздуха, нужной температуры и освещения. Применяя эти методы, физиологи исследуют растения на молекулярном, субклеточном, клеточном и организменном (интактное растение) уровнях. Сейчас в биологических исследованиях широко применяют электронные микроскопы просвечивающего типа с разрешающей способностью 0,15—0,5 нм, в ...
... или транспирация представляют совокупность химических и физических процессов. Чтобы понять механизм физиологического процесса, необходимо выделить его физические и химические компоненты. В связи с этим физиология растений все более нуждается в усовершенствованных методах биохимии. Биохимический подход оказался очень плодотворным в исследованиях таких сложных процессов, как фотосинтез и дыхание. ...
... насекомое тем временем усердно собирает сладкий нектар, предложенный цветком в награду за труды. Цветки правильной формы, то есть имеющие насколько осей симметрии, называются актиноморфными. В классе двудольных растений наблюдается богатое разнообразие цветковых форм. 3. Описание некоторых семейств класса двудольных Семейство лютиковых объединяет около 1500 видов трав и кустарников, многие ...
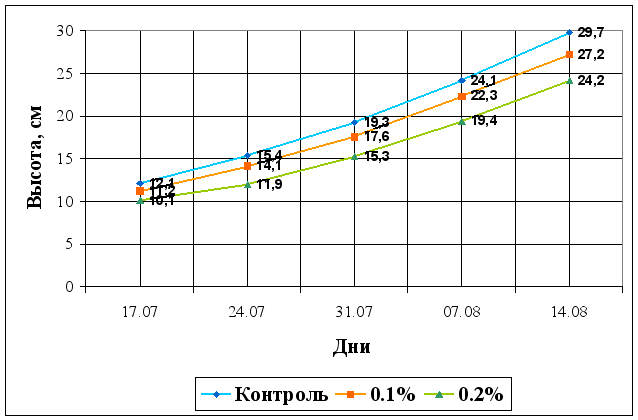
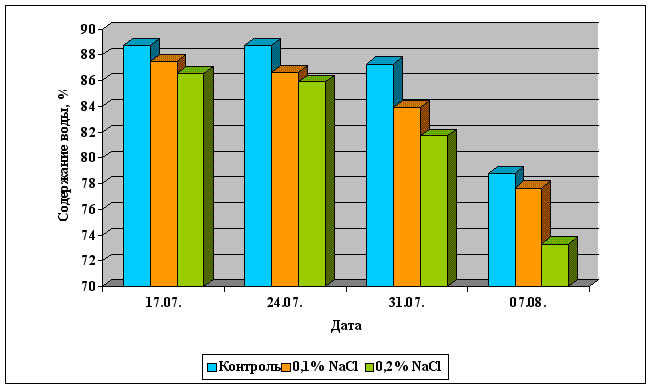
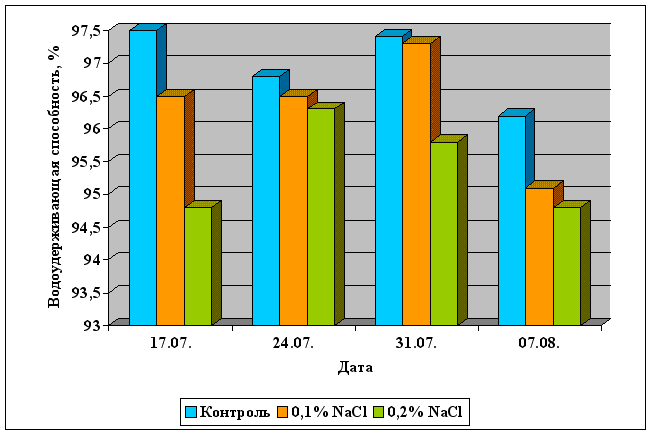
... Областного Университета в период с июня по сентябрь 2003 года. Объектом изучения являлись растения кукурузы сорта Россо. Изучение влияния 6-БАП на рост и биометрические показатели растений кукурузы при разном уровне засоления проводилось в условиях вегетационного опыта (почвенные культуры). Семена кукурузы проращивались при температуре 20оС в термостате в течение 3 дней, а затем высаживались в ...
0 комментариев