Навигация
Становление современных фаунистических областей
53508
знаков
0
таблиц
21
изображение
4.5 Становление современных фаунистических областей
Кайнозойская эра. Несмотря на небольшую продолжительность кайнозойской эры (60-70 млн. лет), ископаемые свидетельства об энтомофауне этого времени наиболее полны.
Изменения климата. Современная физическая картина земной поверхности сформировалась в основном в кайнозойскую эру. Происшедшее в кайнозое похолодание и сильные колебания температуры, последовавшие в четвертичном периоде, оказали большое влияние и на современное распространение насекомых. Эти события приводили к частому и значительному расширению и сужению областей с определенными условиями среды, к смещению климатических поясов к экватору или от него; в мире насекомых они вызывали многократное образование изолированных и реликтовых ареалов. Хотя подобные экстремальные климатические изменения могли происходить и раньше, в пенсильванское время и в начале пермского периода, в мезозое столь сильных колебаний климата не было.
Географические изменения. В кайнозойскую эру продолжалось также отдаление друг от друга и усиление изоляции большинства континентов. Северная Америка и северная часть Европы все еще раздвигались, и к середине эоцена сухопутная связь между ними была прервана. В позднеэоценовое и олигоценовое время по мере продвижения на север континентального блока, включающего Африку, Индию и Средний Восток, теплые прибрежные равнины по южным окраинам Центральной Европы, Центральной Азии и Китая подвергались деформации. Ко времени миоцена Гималайская горная система простиралась почти без разрывов от Европы до южной Азии, и к плиоцену эти горы достигли огромной высоты. С северной стороны этой горной цепи образовался ряд обширных впадин, и на месте многих из них возникли озера неморского происхождения (остатками этих озер являются Черное, Каспийское и Азовское моря). В наше время Гималаи представляют собой климатический и физический барьер между Палеарктикой и Восточной зоогеографической областью.
Австралия и Новая Гвинея тоже двигались на север, заняв положение, близкое к границе с Восточной зоогеографической областью. Резкие контрасты климатических и природных условий в сочетании с относительно недавним передвижением материков ограничивали взаимопроникновение животных и растений Восточной и Австралийской областей.
С юрского периода вплоть до начала плиоцена Северная и Южная Америки были разделены. От Африки и Антарктиды Южная Америка отделилась в конце мелового периода. Панамский перешеек образовался, по-видимому, в середине плиоцена, т. е. 4-6 млн. лет назад. Хотя обмен животными и растениями через этот узкий перешеек шел весьма интенсивно, оба американских континента сохранили ряд четких биотических особенностей. Наибольшие изменения произошли с млекопитающими; североамериканские плацентарные формы в значительной мере вытеснили сумчатых и плацентарных млекопитающих Южной Америки.
Северная Америка и северные районы
Азии на протяжении почти всей кайнозойской эры неоднократно имели связь в области Берингова пролива, и в результате фауны неарктической и палеарктической частей Голарктики стали очень сходны между собой. В Северной Америке значительное увеличение высоты Кордильер и Скалистых гор, происходившее в миоцене, плиоцене и четвертичном периоде, привело к разделению Провинции бассейнов и хребтов (в юго-западной части США) на мелкие, экологически обособленные участки. Провинция бассейнов и хребтов, Скалистые горы и Великие Равнины были подняты на 2-Л км в виде широкого регионального свода, а последующая эрозия придала этому горному ландшафту современный облик. Это поднятие создало «дождевую тень», что привело к распространению луговых сообществ на Великие Равнины. Горы стали холоднее, и на них стало выпадать больше атмосферных осадков.
Среди ископаемых насекомых из верхнеэоценового и нижнеолигоценового балтийского янтаря есть представители нескольких отрядов и подотрядов, которые не встречались в более ранних отложениях: Mantodea, Embioptera и специализированные паразитические жуки Strepsiptera (веерокрылые). Давно известны находки олигоценовых насекомых из северной Европы (Германия и Франция), Северной Америки, Центральной Азии и с Кавказа; они послужили основой для ряда классических исследований. Миоценовые находки известны из северной Европы и с Северного Кавказа, а также из пустыни Мохаве в Калифорнии. Плиоценовых находок значительно меньше.
Большинство ископаемых насекомых, найденных в балтийском янтаре, принадлежат к современным родам. Некоторые семейства высших Hymenoptera и Lepidoptera в коллекциях насекомых из балтийского янтаря отсутствуют и, вероятно, появились в эоцене. Недалеко от Флориссанта (штат Колорадо) в тонких слоях вулканического пепла, осевшего в водоемах у подножия старых Скалистых гор, сохранилось много ископаемых насекомых из олигоцена. Эти насекомые жили в самых разнообразных экологических условиях, от тропических до умеренно-холодных, и их фауна не уступала по многообразию родов современной фауне. Некоторые из этих родов отсутствуют теперь в Северной Америке, но встречаются в других местах - например, мухи цеце (род Glossina) известны сейчас только в Экваториальной Африке. Среди олигоценовых находок из Флориссанта есть интересные златоглазки из семейства Osmylidae, а современные представители этого семейства живут только в тропических областях.
Ледниковые эпохи четвертичного периода (плейстоцен). После плиоцена во всем мире более прохладные эпохи неоднократно чередовались с чуть более теплыми. Во многих районах, особенно в Антарктике и в высоких северных широтах Америки и Евразии, росли и распространялись льды, которые затем таяли. С севера льды продвигались на юг и покрывали обширные территории. Известно четыре главных оледенения, и после каждого из них наступал долгий период с климатом более теплым, чем в настоящее время. Последние два из этих оледенений, а может быть и предыдущие, включали несколько циклов с чередованием ледниковых и межледниковых периодов. Сейчас мы находимся в регрессивной фазе последнего ледникового периода, и лишь остатки северной ледяной шапки сохраняются ныне в виде изолированных ледников или островных масс, как на Гренландии.
Расселение, дизъюнктивные ареалы и видообразование. Изменения климата во время смены ледниковых и межледниковых периодов оказали огромное влияние на живую природу Северной Америки, особенно в северных районах, отличавшихся умеренным климатом. В то время как обмен между фаунами Северной Америки и западной Европы отсутствовал, северо-западная часть Северной Америки и северо-восточная часть Азии были связаны по суше Беринговым мостом, который мог служить путем для расселения животных. В результате, когда условия на севере стали такими же, как в наше время, или даже более теплыми, многие виды насекомых, приспособленные к умеренно-холодному климату, расселились по всей Голарктике, распространяясь из Северной Америки в Азию или в обратном направлении. Во время следующего похолодания, при новом увеличении площади ледников ареалы этих видов расширялись к югу, так как южные окраины их ареалов становились холоднее и лучше соответствовали потребностям этих насекомых. Когда северные границы ареалов смещались к югу от Берингова моста, ареал каждого вида распадался на две части, между которыми не было генетического обмена. Если такое разобщение было достаточно долгим, популяции внутри каждой из этих частей могли стать самостоятельными видами. Когда ледники таяли и наступал очередной теплый межледниковый период, ареалы видов перемещались к северу. При этом вновь происходило смешивание азиатских и североамериканских видов, многие из которых были близкородственными, но уже генетически изолированными; в результате формировалась новая группа голарктических видов, а затем весь этот длительный процесс образования новых видов под влиянием климатических факторов повторялся снова.
Такой же механизм лежал в основе видообразования в горных районах в пределах одного континента. Например, холодолюбивый род веснянок Allocapnia, распространение которого ограничено холодными ручьями восточной части Северной Америки, дал начало большой группе видов (около 38); этому способствовали прежде всего волны рассеяния, связанные с флуктуациями климата, а затем изоляция географически обособившихся популяций родительских видов в нескольких горных районах-на севере и юге Аппалачских гор, на плато Камберленд и в горах Озарк-Уошито (штаты Миссури, Арканзас и Оклахома). Судя по имеющимся данным, все или почти все эти виды возникли, вероятно, в плейстоцене, т.е. в последние 3 млн. лет.
Ареал северного холодолюбивого вида ручейников Glossosoma intermedium в целом сейчас простирается от западной Европы через Азию на восток до Миннесоты и Миссури. На протяжении всего огромного ареала вид поразительно однороден, и это указывает на то, что он распространился по всей Голарктике во время последнего оледенения - возможно, всего лишь 12-30 тыс. лет назад. Однако ареал этого вида разделен на ряд изолированных популяций, что создает условия для образования новых видов в будущем.
В юго-западных районах США благодаря климатическим изменениям в четвертичном периоде возник еще один механизм видообразования. Ледниковые периоды были холодными и дождливыми, а межледниковые - жаркими и сухими. Целый ряд насекомых в этих районах, в том числе многие жуки, осы и пчелы, приспособлен к условиям пустыни. В дождливые эпохи их ареалы были ограничены небольшими изолированными слегка приподнятыми пустынными или полупустынными районами, расположенными между холодными, влажными горными хребтами. В это время такие изолированные, приспособленные к сухому климату популяции становились видами. В следующий межледниковый период сухие жаркие районы расширялись за счет прохладныхи влажных, в результате возникавших контактов происходило расселение и смешение видов.
Благодаря находкам в торфе и богатом органическим веществом иле появляется все больше данных об ископаемых насекомых четвертичного периода. У некоторых насекомых, например у жуков, части экзоскелета настолько прочны, что их можно вымывать из рыхлых отложений, не повреждая. Недавно были изучены находки Coleoptera из позднеплейстоценовых (висконсинских) отложений. Большинство этих ископаемых жуков принадлежит к современным видам жуков из семейств, приспособленных к умеренному климату и, судя по полученным данным, эволюционирующих сравнительно медленно. Перемещения, сужения и расширения климатических поясов в четвертичном периоде имели для насекомых многосторонние последствия, важнейшими из которых были изменения их ареалов и характера расселения ; при этом темпы эволюции не были во всех группах одинаковыми.
4.5 Древние реликтовые фауныСреди современных насекомых, живущих в областях, входивших в древнюю Гондвану, огромное число форм принадлежит к семействам, известным только из Австралии, Новой Зеландии и Южной Америки. Некоторые другие встречаются, помимо указанных районов, также в Южной Африке. Большинство этих насекомых приспособлены к южной умеренной зоне, ведут оседлый образ жизни и не имеют средств для преодоления океанических пространств. К таким насекомым относятся, например, нелетающие растительноядные клопы из семейства Peloridiidae (Hemiptera), долгоносики из семейства Belidae, нелетающие сверчки из семейства Cylindrachetidae, три семейства и одно подсемейство веснянок (Plecoptera). В этот южный фаунистический комплекс входят также примитивные скорпионовые мухи из семейства Nannochoristidae - единственные дожившие до наших дней представители отряда Mecoptera, имеющие холодолюбивых водных личинок. Это семейство известно и из пермских отложений Австралии. Из двукрылых представителями этой фауны являются род Pelecorhynchus, родственный толстоножкам (сем. Bibionidae) и слепням (сем. Tabanidae), род Trichophthalma (сем. Nemestrinidae), подсемейство Ceratomerinae мух-толкунчиков (сем. Empididae) и примитивное семейство комаров Blephariceridae. Есть и другие примеры, указывающие на общность происхождения этой фауны. Большинство таких видов - дожившие до наших дней представители древних примитивных ветвей, и это заставляет предполагать, что когда-то до раскола Гондваны у них были сплошные обширные ареалы. В каком-то смысле эти насекомые-реликтовые, эндемичные и дизъюнктивные в одно и то же время!
ВЫВОДЫ
Существует несколько гипотез о происхождении насекомых и их взаимоотношениях с другими классами членистоногих. Основные затруднения заключаются в том, что палеонтологи еще не нашли отпечатков ископаемых форм, которые могли бы с достаточным основанием считаться предками насекомых. Тем не менее уже накопленные материалы по ископаемым насекомым позволяют относить время их возникновения к началу девонского геологического периода и в самых общих чертах представить себе существовавшую тогда экологическую обстановку. В девонский период, начавшийся 375 млн. лет назад и продолжавшийся 50 млн. лет, осуществлялся важнейший эволюционный этап развития жизни — выход растений из водоемов на сушу. К началу этого периода на суше растений не было, а жизнь концентрировалась в водоемах, где имелись водоросли и беспозвоночные.
В настоящее время полагают, что в дополнение к многочисленным современным отрядам насекомых прежде, судя по ископаемым остаткам, существовало еще 11 или даже больше отрядов. Большинство исчезнувших отрядов впервые появились в отложениях, относящихся к Пенсильванию или ранней перми, и в основном вымерли уже к концу пермского периода. Среди этих исчезнувших форм были и ранние предки некоторых современных отрядов насекомых. Большинство этих ископаемых предков морфологически были более примитивны и менее специализированы, чем их потомки; поэтому в позднем палеозое различия между группами насекомых еще не были такими четкими, как между соответствующими современными отрядами. С другой стороны, изучение поздних палеозойских насекомых ясно показывает, что к концу Пенсильвания основные этапы эволюции насекомых были уже завершены. Это относится к развитию крыльев, их складыванию, дифференциации способов питания и выработке жизненных циклов с полным превращением. Ниже даются краткие описания тех вымерших отрядов насекомых, которые уже достаточно изучены.
Морфологическая организация, которая характерна для насекомых, обеспечила им фундаментальный морфологический прогресс (ароморфоз). На основе этой организации они освоили воздушно-наземную среду обитания. В дальнейшем насекомые приспособились к самым разнообразным частным средам, в результате чего у них возникли различные конкретные приспособительные адаптации (идиоадаптации). На основе этих адаптации в прошлом и в настоящее время у насекомых происходит интенсивное видообразование.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1) Андрианова Н.С. Экология насекомых: Курс лекций. – М., 1970. – 158 с.
2) Бей-Биенко Г. Я. Общая энтомология. Издание третье, дополненное. М.:Высшая школа, 1980. - 416 с.
3) Грант В. Эволюция организмов. - М., 1980.
4) Жизнь животных: В 6-ти томах. – М., 1985.
5) Майр Э. Популяции, виды и эволюция. М., 1974.
6) Мамаев Б.М. Энтомология. - М.: Мир, 1976. – 426 с.
7) Росс Г. и др. Энтомология: пер. с англ. – М.: Мир, 1985. – 576 с.
8) Савчук М.П. Зоологія безхребетних. – К., 1965. – 503 с.
9) Северцов А.Н. Эволюционная теория. – М., 1967.
10) Щербак Г.Й. Зоологія безхребетних. Кн. 1-3. – К.: Либідь, 1995.
11) Яблоков А.В., Юсупов А.Г. Эволюционное учение. – М.: Высш. шк., 1989. – 335 с.
Похожие работы
... не существует. Однако и синтетическая теория эволюции имеет ряд трудностей, что ставит эволюционистов в трудное положение, и на которых основываются недарвиновские концепции эволюции. Доказательства эволюции живой природы Эволюционные процессы наблюдаются как в естественных условиях, так и в лаборатории. Известны случаи образования новых видов. Также описаны случаи развития новых свойств ...
... насекомых, и там будут развиваться совершенно другие виды. При этом стоит отметить, что многие насекомые проявляют четкую зависимость от вида гриба, разлагающего древесину. Таблица 1 Распределение дереворазрушающих грибов подзоны средней тайги Республики Коми по трофическим группам (по Косолапову, 2008) Трофические группы Число видов Сапротрофы На валежной древесине 318 ...
... отбора" (1859 г.) этой формулировки нет. Даже спустя 12 лет, в работе "Происхождение человека и половой отбор", осмотрительный Дарвин пишет лишь об эволюции вообще, совершенно не касаясь "промежуточного звена". О происхождении же человека от обезьяны заявили в 1863 г. дарвинисты Фохт, Гексли и Геккель. Именно Геккель говорил о "недостающем звене" между высшими приматами и человеком, имея в виду ...
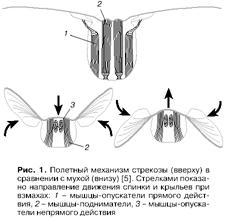
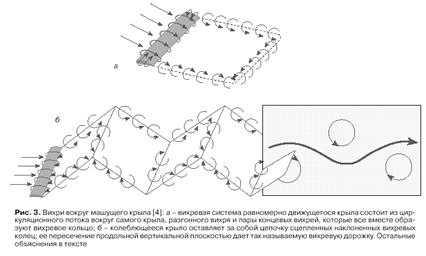
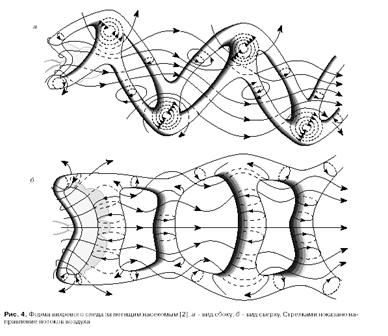
... противоположен по направлению момент сил, переданному машущими крыльями окружающей среде. Как следствие взаимодействия машущих крыльев с воздухом за летящим насекомым остается аэродинамический след, структура которого содержит информацию о природе сил, создаваемых в машущем полете. Как образуются вихревые кольца. Рассмотрим работу машущего крыла с позиции принципов, заложенных в обоих подходах. ...
0 комментариев