Навигация
Регуляция экспрессии генов на уровне транскрипции у прокариот
40945
знаков
0
таблиц
1
изображение
2. Регуляция экспрессии генов на уровне транскрипции у прокариот
Регуляция транскрипции в клетках осуществляется на уровне индивидуальных генов, их блоков и даже целых хромосом. Возможность управления многими генами, как правило, обеспечивается наличием у них общих регуляторных последовательностей нуклеотидов, с которыми взаимодействуют однотипные факторы транскрипции. В ответ на действие специфических эффекторов такие факторы приобретают способность с высокой точностью связываться с регуляторными последовательностями генов. Следствием этого является ослабление или усиление транскрипции (репрессия или активация) соответствующих генов. Три основных этапа транскрипции – инициация, элонгация и терминация, рассмотренные нами выше, используются бактериальными клетками для регуляции синтеза РНК. То же, по-видимому, характерно и для остальных живых организмов. Ниже приведены примеры регуляторных воздействий на транскрипцию.
Регуляция на уровне инициации транскрипции
Активность многих генов прокариот регулируется с помощью белковых факторов, взаимодействующих с регуляторными участками промоторов генов. При этом происходят как активация транскрипции генов, так и подавление считывания генетической информации РНК-полимеразами. В первом случае регуляторные белковые факторы называют активаторами, осуществляющими позитивную регуляцию транскрипции, а во втором – репрессорами. Регуляцию, связанную с подавлением транскрипции, называют негативной. Механизмы, при помощи которых активаторы стимулируют инициацию транскрипции, могут быть рассмотрены с двух точек зрения – кинетической и структурной. Поскольку активация промоторов путем образования открытых комплексов является лимитирующей стадией на пути активации транскрипции в целом, действие различных активаторов может быть охарактеризовано по изменению (увеличению) значений кинетических параметров реакций, происходящих на разных этапах активации. Так, при действии активирующего комплекса Crp–cAMP на lac-промотор происходит десятикратное увеличение равновесной константы ассоциации KB РНК-полимеразы с промотором с образованием закрытого комплекса. Активация промотора PRM фага λ, опосредованная cI-белком (см. ниже), характеризуется пяти–десятикратным увеличением константы скорости kf перехода закрытых комплексов в открытые. Активирующее действие белка λ-cII на промотор РRE сопровождается изменением обоих вышеупомянутых кинетических параметров. Активация транскрипции может быть также опосредована увеличением скорости освобождения промотора РНК-полимеразой после инициации синтеза РНК. Многие активаторы транскрипции, в том числе и Crp–cAMP, сгибают молекулу ДНК после взаимодействия с ней, причем центр такого изгиба находится в сайте связывания активатора. Однако с использованием мутантных белков было установлено, что изгибание ДНК и связывание активаторов с ДНК как таковое еще не обеспечивают активацию транскрипции. В большинстве случаев абсолютно необходимым условием активации является наличие контакта между специфическими областями поверхностей молекул активатора и РНК-полимеразы, часто с ее α-субъединицами. Важным следствием образования контактов между активаторами и холоферментом РНК-полимеразы является часто наблюдаемый синергизм в связывании обоих белков с соответствующими промоторами. При этом мутации в сайтах связывания активаторов или промоторе как таковом могут предотвращать активацию транскрипции путем изменения конформации молекулы связанного активатора или контактного участка на РНК-полимеразе. Последовательности нуклеотидов промоторных участков генов, с которыми взаимодействуют молекулы репрессора, получили название операторов. Во многих случаях репрессор связывается с оператором только в присутствии низкомолекулярного лиганда, специфически взаимодействующего с репрессором. Такие низкомолекулярные эффекторы получили название корепрессоров. Они часто требуются и для функционирования белков- активаторов транскрипции. Простейший механизм репрессии заключается встерическом блокировании связывания РНК-полимеразы с промотором. Это происходит в том случае, если последовательности нуклеотидов мест посадки РНК-полимеразы на промотор и репрессора на оператор перекрываются. Некоторые бактериальные белки-репрессоры могут оказывать свое негативное действие на этапы инициации, происходящие после связывания РНК-полимеразы с промотором. Например, молекулы репрессора gal- оперона E. coli, связавшиеся с операторами OE и OI, центры последовательностей которых расположены соответственно на расстояниях –60,5 и +53,5 по отношению к точке инициации транскрипции, вызывают образование петли участка ДНК, заключенного между ними, но не препятствуют взаимодействию РНК-полимеразы с промотором. Они оказывают свое действие на последующие этапы инициации, предшествующие образованию первой фосфодиэфирной связи. В том случае, если лишь одна молекула репрессора связывается с внешним оператором OE, он частично ингибирует транскрипцию путем взаимодействия с α-субъединицей РНК-полимеразы. Это сопровождается понижением уровня, но не полным прекращением синтеза РНК gal-оперона, т.е. более тонким изменением уровня экспрессии соответствующих генов. Распространенным механизмом активации транскрипции с помощью белков-активаторов является облегчение ее инициации РНК-полимеразой после образования контакта между ферментом и белком-активатором, связанными с регуляторной областью промотора, что сопровождается конформационными изменениями РНК-полимеразы. У бактерий имеются белки- регуляторы, обладающие активностью как репрессора, так и активатора транскрипции. Такими "амфотерными" свойствами обладает, в частности, репрессор cI фага λ. Белок-активатор катаболитных оперонов (Crp-белок) активирует транскрипцию бактериальных генов, продукты которых участвуют в расщеплении (катаболизме) различных органических соединений (преимущественно сахаров), используемых растущей бактериальной клеткой в качестве источника углерода. Свои свойства активатора Crp-белок приобретает лишь в комплексе с циклическим AMP (сAMP). Внутриклеточная концентрация сAMP возрастает у бактерий, растущих на бедных питательных средах, и понижается в условиях избытка легко усвояемых источников углерода, например глюкозы. Поэтому система Crp–сAMP обеспечивает включение экспрессии катаболитных оперонов лишь на бедных питательных средах. Crp- белок может выступать и в роли репрессора транскрипции генов галактозного оперона E. coli. Если все гены катаболитных оперонов активируются Crp-белком в присутствии сAMP, то негативная регуляция их транскрипции происходит индивидуально. Хорошо известными примерами такого рода являются регуляции транскрипции lac-оперона E. coli под действием Lac- репрессора, а также галактозного и арабинозного оперонов специфическими белками-репрессорами этих оперонов. Низкомолекулярные эффекторы могут изменять активность РНК-полимеразы не только опосредованно через белки-регуляторы, но и непосредственно при взаимодействии с ферментом. С помощью гуанозинтетрафосфата (ppGpp) в клетках E. coli осуществляется координация экспрессии генов рибосомных РНК (рРНК) и белков. Этот необычный нуклеотид (известный также под названием "магического пятна") синтезируется бактериальными клетками в условиях внутриклеточного недостатка аминокислот, что приводит к значительному снижению интенсивности транскрипции генов рРНК и белков и одновременной стимуляции синтеза РНК оперонов, контролирующих биосинтез аминокислот. В присутствии ppGpp очищенная РНК-полимераза прекращает синтез рРНК с одного из двух промоторов этих оперонов, что приводит к ослаблению, но не полному прекращению их транскрипции. Известны мутации, локализованные в гене β-субъединицы РНК-полимеразы E. coli, приводящие к прекращению влияния ppGpp на синтез РНК, что подтверждает наличие непосредственного контакта между нуклеотидом и ферментом в процессе транскрипции. Некоторые регуляторные элементы бактерий, участвующие в активации транскрипции, так же как и энхансеры эукариот, могут располагаться на большом расстоянии (нескольких сотен нуклеотидов) от промоторов, на которые они оказывают свое действие. В этом случае контакт активатора с РНК-полимеразой обеспечивается благодаря выпетливанию участка ДНК, расположенного между данными регуляторными элементами, что приводит к пространственному сближению двух белков. Прямое доказательство образования таких петель на ДНК E. coli было, в частности получено с помощью электронной микроскопии для белков NtrC и NifA, действующих соответственно на промоторы генов glnA и nifH. Другим путем достижения белком-активатором молекулы РНК-полимеразы на удаленном промоторе (например промоторе поздних генов бактериофага Т4) является его перемещение вдоль отрезка ДНК, разделяющего эти два регуляторных элемента. Процесс такого перемещения может быть инициирован последовательностями нуклеотидов, расположенными выше или ниже промотора на расстоянии нескольких сотен пар оснований. Одним из давно обсуждающихся вопросов является необходимость изменения структуры ДНК в окрестностях промоторов под действием белков- активаторов для активации транскрипции. В ряде случаев такие доказательства были получены. Так, в mer-локусе E. coli, обеспечивающем устойчивость бактериальных клеток к ионам ртути, связывание Mer-белка с регуляторным участком промотора (merT) в присутствии ртути сопровождается раскручиванием спирали ДНК в районе промотора на 50о. Это приводит к образованию правильного расстояния между сайтами связывания активатора и промотором, так как первый расположен необычно – между нуклеотидами в положениях –35 и –10 промотора. Без такого изменения структуры ДНК связавшийся с ним активатор не может образовать правильного контакта с РНК-полимеразой.
Похожие работы
... в различных температурных условиях. Эта корреляция не распространялась на растения, подвергнутые действию недостатка питательных веществ или засухи. При изучении влияния низко температуры на экспрессию генов в Poncirus trifoliata из закаленных к холоду растений были клонированы шесть кДНК, представляющие уникальные индуцируемые холодом последовательности. В ходе экспериментов было обнаружено, ...
... » в транскрибируемой последовательности, который может остановить транскрипцию гена. Модификация и / или процессинг, которые могут предшествовать образованию зрелой функциональной РНК, также регулируются. Экспрессия генов может регулироваться и на уровне трансляции мРНК с образованием белков. И в этом случае специфическая регуляция, как правило, осуществляется на начальном этапе декодирования. ...
... генов-"мишеней". Эти системы можно рассматривать как некое подобие генных сетей, описанных выше. Хромосомный контроль развития Вышеизложенные представления о генном контроле развития не исключают других уровней контроля, в частности, хромосомный. Важность этого уровня регуляции можно проиллюстрировать примерами, показывающими, что вне хромосомного (хроматинового) контекста невозможно ...
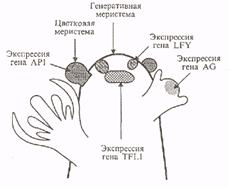
... этапах развития соцветия и у мутантов, культивируемых при длинном дне. Карпелоидные структуры у мутантов По также изредка образуются только у очень старых растений. 1.3 Экспрессия генов идентичности цветковых меристем Хотя экспрессия генов FLO/LFY и SQUA/AP1 может быть индуцирована независимо друг от друга, очевидно, что наличие активности только одного из этих генов недостаточно для развития ...
0 комментариев