Навигация
3. Трансляция у эукариот
Бактерии обладают единственной универсальной системой трансляции, основные механизмы функционирования которой были кратко рассмотрены выше. В отличие от этого, клетки животных кроме основной системы трансляции, локализованной в цитоплазме, имеют дополнительную систему трансляции митохондрий, которая по ряду свойств приближается к бактериальной. Клетки растений обладают еще одной дополнительной системой биосинтеза белка, функционирующей в хлоропластах. Большинство данных о механизмах биосинтеза белка у эукариот было получено с использованием бесклеточных белоксинтезирующих систем. В последнее время важные результаты о механизмах трансляции у эукариот были получены с использованием стабильно трансформированных клеток животных и растений, выращиваемых в культуре. В ходе этих исследований установлено, что у растений и животных в основном функционируют одни и те же механизмы трансляции. Инициация биосинтеза белка эукариотическими рибосомами Как будет видно из дальнейшего изложения, инициация трансляции эукариотических мРНК может осуществляться, по крайней мере, тремя способами. В соответствии с первым наиболее распространенным механизмом (модель сканирования) рибосомы после взаимодействия с 5'-концевой последовательностью мРНК осуществляют поиск инициирующего AUG-кодона, перемещаясь вдоль 5'UTR. При реализации второго механизма рибосомы инициируют биосинтез белка на внутренних AUG-кодонах, удаленных от 5'-концевой кэп-группы. И, наконец, после освобождения полипептида из транслирующего комплекса рибосомы, не отделяясь от мРНК, способны реинициировать биосинтез белка на следующем инициирующем кодоне.
Факторы инициации трансляции. Большинство молекулярных механизмов, осуществляющих регуляцию экспрессии генов на уровне трансляции, реализуется на стадии инициации биосинтеза белка. По-видимому, этот факт находит свое отражение в большой сложности аппарата инициации трансляции. Помимо субъединиц эукариотических рибосом и белков, обычно ассоциированных с 5'- и 3'-концевыми последовательностями мРНК, в инициации принимают участие по меньшей мере 11 белковых факторов, построенных более чем из 25 полипептидов Элонгация полипептидных цепей в ходе эукариотической трансляции традиционно пользовалась меньшим вниманием исследователей по сравнению с инициацией, поскольку считалось, что ее механизмы в основных чертах идентичны таковым бактерий. Дальнейшие исследования показали, что данная точка зрения в основном соответствует действительности, хотя эукариотическая система трансляции обладает более сложным набором факторов элонгации.
Факторы и механизмы элонгации. Эукариотические клетки содержат в большом количестве фактор элонгации eEF1A, который является функциональным гомологом бактериального фактора EF1A(EF-Tu). Так же как и у бактерий, этот фактор образует тройной комплекс с GTP и аминоацил-тРНК, обеспечивая вхождение последней в А-участок элонгирующей рибосомы. Два других эукариотических фактора eEF1B и eEF2 резко отличаются от бактериальных функциональных аналогов EF1B(EF-Ts) и EF2(EF-G) по аминокислотным последовательностям. Гетеротримерный фактор eEF1B, как и его бактериальный аналог, катализирует обмен GDP на GTP в комплексе eEF1A–GDP. Фактор eEF2, по аналогии с бактериальными системами, обеспечивает транслокацию пептидил-тРНК в P-участок рибосом и перенос деацилированной тРНК в E-участок. У высших организмов этот фактор служит мишенью регуляторных воздействий через фосфорилирование. Замечательным свойством факторов eEF1A и eEF2 является способность связываться с компонентами цитоскелета эукариотических клеток. Полагают, что это их свойство может обеспечивать один из механизмов внутриклеточного транспорта мРНК, направляющих ее в полисомы. Растущий полипептид выводится в цитоплазму через канал, начало которого расположено на поверхности рибосомы, где он взаимодействует с белками, распознающими сигнальную последовательность, или с другими цитоплазматическими факторами, которые обеспечивают его направленный транспорт внутри эукариотической клетки. У бактерий растущая полипептидная цепь может вызывать уменьшение скорости элонгации, а природа предпоследней аминокислоты оказывает сильное влияние на терминацию трансляции. Предполагают, что такого рода эффекты являются следствием взаимодействия между строящимся пептидом и факторами трансляции, рРНК или непосредственно каналом, через который он переносится к поверхности рибосомы. Подобные механизмы, по-видимому, функционируют и у эукариот. У дрожжей, как и у E. coli, скорость элонгации трансляции снижается в присутствии редко встречающихся кодонов в мРНК. Наличие определенного числа таких кодонов вблизи сайта инициации трансляции значительно снижает скорость считывания соответствующих ОРС. На скорость декодирования мРНК рибосомами оказывают влияние и характер фолдинга строящихся полипептидных цепей, а также сигнальные последовательности аминокислот, определяющие направление их посттрансляционного транспорта внутри эукариотических клеток.
Терминация трансляции
В эукариотических белоксинтезирующих системах терминация трансляции, как и у бактерий, контролируется специфическими рилизинг-факторами. Однако у эукариот эти факторы менее разнообразны. В частности, у них отсутствует функциональный аналог бактериального фактора RRF/RF4.
Факторы терминации. По современным представлениям, элонгирующая эукариотическая рибосома распознает стоп-кодоны, находящиеся в одной рамке с основными ОРС, после взаимодействия с гетеродимерным комплексом рилизинг-факторов (RF), в состав которого входят факторы eRF1 и eRF3. Фактор eRF1 необходим для распознавания всех трех терминирующих кодонов (UAA, UAG и UGA) и освобождения синтезированного полипептида. Фактор eRF3 является GTPазой, обладающей гомологией с eEF1A, которая, гидролизуя GTP, стимулирует терминацию независимо от последовательности нуклеотидов в терминирующих кодонах.
Заключение
Определение функций продуктов новых генов на основе их первичной структуры осложняется тем, что многие белки и нуклеиновые кислоты проявляют свою активность и функционируют только в составе больших надмолекулярных комплексов, размеры которых часто приближаются к размерам рибосом. При этом многие белки сами по себе не обладают ферментативной активностью, а выполняют вспомогательные функции, например, молекул-адаптеров, обеспечивающих сборку комплексов и создающих молекулярные интерфейсы для их взаимодействия с регуляторными и каталитическими субъединицами. Наличие таких комплексов, как это было установлено в последнее десятилетие, особенно характерно для клеток эукариотических организмов. Так, исследование молекулярных механизмов транскрипции у эукариот привело к развитию представления о транскриптосоме, гигантскому белковому комплексу, в который кроме холофермента РНК-полимеразы с ее многочисленными субъединицами входят факторы транскрипции, белки-адаптеры, белковые компоненты системы репарации и т.п. При этом размер транскриптосомы приближается к таковому целых рибосом. В гигантские надмолекулярные комплексы организованы и молекулярные машины системы синтеза ДНК (реплисомы), процессинга и редактирования РНК (сплайсомы и эдитосомы), молекулярные компоненты системы протеолитической деградации белков (протеасомы). Создается впечатление, что организация генетических систем, функционирующих на основных этапах реализации генетической информации, в гигантские пространственно упорядоченные комплексы, является общебиологическим принципом.
Список использованной литературы
1. Патрушев Л.И. Экспрессия генов. – М.: Наука, 2000. – 800 с., ил.
2. Айала Ф., Кайдегер Дж.. Современная генетика: т.2. М.: Мир, 1988.
3. Биология: в 2 кн. Кн. 1: Жизнь. Гены. Клетка. Онтогенез. Человек./ под ред. В.Н.Ярыгина.
4. Биология: Большой энциклопедический словарь/ гл. ред. М.С.Гиляров.
5. Вилли К., Детье В.. Биология (биологические процессы и законы). М.:Мир, 1975.
Похожие работы
... в различных температурных условиях. Эта корреляция не распространялась на растения, подвергнутые действию недостатка питательных веществ или засухи. При изучении влияния низко температуры на экспрессию генов в Poncirus trifoliata из закаленных к холоду растений были клонированы шесть кДНК, представляющие уникальные индуцируемые холодом последовательности. В ходе экспериментов было обнаружено, ...
... » в транскрибируемой последовательности, который может остановить транскрипцию гена. Модификация и / или процессинг, которые могут предшествовать образованию зрелой функциональной РНК, также регулируются. Экспрессия генов может регулироваться и на уровне трансляции мРНК с образованием белков. И в этом случае специфическая регуляция, как правило, осуществляется на начальном этапе декодирования. ...
... генов-"мишеней". Эти системы можно рассматривать как некое подобие генных сетей, описанных выше. Хромосомный контроль развития Вышеизложенные представления о генном контроле развития не исключают других уровней контроля, в частности, хромосомный. Важность этого уровня регуляции можно проиллюстрировать примерами, показывающими, что вне хромосомного (хроматинового) контекста невозможно ...
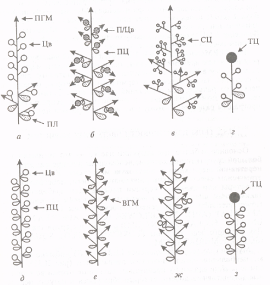
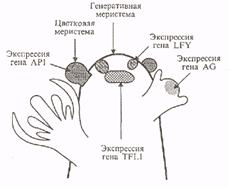
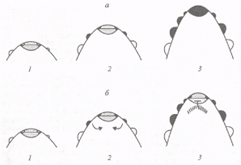
... этапах развития соцветия и у мутантов, культивируемых при длинном дне. Карпелоидные структуры у мутантов По также изредка образуются только у очень старых растений. 1.3 Экспрессия генов идентичности цветковых меристем Хотя экспрессия генов FLO/LFY и SQUA/AP1 может быть индуцирована независимо друг от друга, очевидно, что наличие активности только одного из этих генов недостаточно для развития ...
0 комментариев