Навигация
Катаболизм и стресс у растений
39242
знака
0
таблиц
3
изображения
МИНИСТЕРСТВО НАУКИ И ОБРАЗОВАНИЯ РОССИЙСКОЙ
ФЕДЕРАЦИИ
ГОУ ВПО «НОВОСИБИРСКИЙ ГОСУДАРСТВЕННЫЙ
ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ»
ИНСТИТУТ ЕСТЕСТВЕННЫХ И
СОЦИАЛЬНО-ЭКОНОМИЧЕСКИХ НАУК
КАФЕДРА БОТАНИКИ И ЭКОЛОГИИ
РЕФЕРАТ ПО ФИЗИОЛОГИИ РАСТЕНИЙ
Тема: Катаболизм и стресс у растений
Выполнил:
Калиниченко Н.А.
320 группа
Проверила: д.б.н., профессор
Барахтенова Л.А.
Новосибирск 2007
Содержание:
1. Введение...............................................................................................................3
2. Значение и функции катаболизма для растения...............................................4
3. Биогенный стресс и катаболизм.......................................................................12
4. Заключение.........................................................................................................20
5. Список литературы............................................................................................25
1. ВВЕДЕНИЕ
"В организмах совершается не только процесс создания, т.е. питания и роста, но, рука об руку с ним, идет процесс разрушения и выделения".
"Организм не ассимилирующий теряет в весе, - этим обнаруживается его разрушение; но оно идет и одновременно с ассимиляцией, о чем мы судим по продуктам выделения. Без этого процесса разрушения невозможны проявления жизни. Жизненная деятельность находится, разумеется, в известных пределах, в прямом отношении с этим процессом разрушения. Чем энергичнее трата, тем энергичнее жизненная деятельность. Только совокупность двух процессов, созидания и разрушения, характеризует живое тело. Организм живет только пока разрушается".
К.А. Тимирязев
Обмен веществ слагается из множества одновременно протекающих реакций, среди которых можно выделить два важнейших противоположно направленных процесса: анаболизм и катаболизм - синтез и деструкция. В данном реферате мы рассмотрим различные аспекты катаболитических процессов наблюдаемых у растений при стрессе.
Большинство современных исследований направлено на изучение синтетических процессов, что определяется не только их очевидной важностью, но и появлением и совершенствованием эффективных методов исследования, таких, как радиоактивные индикаторы, хроматография, электрофорез и т.д.
Изучение катаболизма биополимеров и липидов сопряжено с гораздо большими трудностями, чем их синтеза. Например, если поставлена задача изучения метаболизма определенного белка, то его необходимо "подметить" с помощью того или иного изотопа, ввести белок в заданный компартмент клетки и затем прослеживать за особенностями его превращения.
Проведение всех трех операций связано со значительными трудностями. Это главным образом и предопределяет неизмеримо меньшее число публикаций, посвященных процессам катаболизма биополимеров и липидов.
Другая причина - недооценка функциональной роли процессов деградации сложных соединений. Сейчас существует мало публикаций где реакции клеточного метаболизма представлены в виде трех функциональных блоков - катаболизма, анаболизма, а также роста и дифференцировки. [2, стр.8].
2. ЗНАЧЕНИЕ И ФУНКЦИИ КАТАБОЛИЗМА ДЛЯ РАСТЕНИЯ
Можно говорить о множественной роли процессов катаболизма.
Во-первых, роль корректирующего фактора (рис. 1). Он обеспечивает, устранение биополимеров с «некорректной» структурой, вызванной тремя возможными причинами: ошибками в ходе образования этих соединений; большей или меньшей потерей их активности в результате последующей модификации; таким изменением окружающей среды, при котором эти биополимеры или липиды не могут в полной мере выполнять свои функции [2, стр.18].
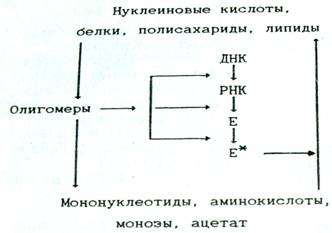
Рис. 1. Схема взаимосвязи катаболизма и анаболизма, отражающая субстратную и регуляторную функции катаболизма
Во-вторых, субстратная роль. Процессы катаболизма обеспечивают синтез биополимеров мономерными субстратами, а синтез липидов - ацетатом, что особенно важно в условиях недостаточного насыщения клеток этими веществами.
В-третьих, энергетическая роль. При дальнейшей деградации мономерных продуктов и ацетата и вовлечении их в процессы дыхания образуются АТФ и НАД(Ф)Н, обеспечивающие протекание анаболических реакций.
В-четвертых, сигнальная функция. Известно, что особенности структуры нуклеиновых кислот, белков и некоторых поли- и олигосахаридов (в том числе входящих в состав гликопротеидов и гликолипидов) используются в клетках для хранения, передачи и реализации информации [2, стр.19].
В процессе деградации биополимеров происходит постепенное уменьшение заключенного в этих молекулах объема информации. Однако трудно себе представить, что в ходе эволюции не были использованы информационные свойства фрагментов молекул биополимеров, образующихся при деструкционном каскаде в результате действия эндо- и экзогидролаз. Исследования в этом направлении показали, что, во-первых, частичная деградация предобразованных молекул является широко распространенным способом формирования функциональных молекул нуклеиновых кислот и белков в реакциях соответственно посттранскрипционного процессинга РНК и посттрансляционной модификации белков.
Во-вторых, накапливается все больше информации о том, что некоторые олигонуклеотиды, олигопептиды и олигосахариды являющиеся продуктами катаболизма соответствующих полимеров обладав свойствами гормонов или активаторов и ингибиторов различных процессов метаболизма и в конечном итоге оказывают влияние на рост и морфогенез растений.
Можно считать, что сигнальные свойства олигомерных промежуточных продуктов катаболизма реализуются путем воздействия на транскрипцию, трансляцию или на предобразованные ферменты связанные с процессами анаболизма, а также на мембранные структуры (см. рис. 1)
Приведенные выше рассуждения о сигнальных свойствах олигомерных производных биополимеров справедливы и в отношении промежуточных продуктов деградации липидов, в состав которых входят полиеновые (ненасыщенные) жирные кислоты — линолевая, линоленовая, а у низших грибов и бактерий - арахидоновая и эйкоэапентаеновая, обладающие соответственно двумя, тремя, четырьмя и пятью двойными связями. Сигнальные свойства полиеновых жирных кислот определяются реакциями оксигенирования молекул по местам двойных связей и последующими реакциями, в результате осуществления которых появляется множество производных жирных кислот. Некоторые из этих продуктов оксигенирования являются физиологически активными веществами.
Деградация мембранных липидов может приводить к драматическим последствиям, в связи с влиянием на физико-химические свойства мембран и условия функционирования катализирующих разнообразные процессы мембранных белков-ферментов, транслокаторных систем, рецепторов и т.д.
Скорость деградации и окислительных реакций в мембранных липидах имеет большое значение для регуляторной и информационной роли мембран в клеточном метаболизме [9, стр.557].
Некоторые из промежуточных продуктов метаболизма биополимеров и липидов в ходе эволюции животных и растительных организмов стали выполнять функции гормонов - физиологически активных веществ, вырабатываемых в одних органах, транспортируемых в другие и изменяющих в них интенсивность и направленность метаболизма и в конечном итоге влияющих на реализацию программ роста, морфогенеза, устойчивости и др.
Принципиальное отличие этих систем у растительных и животных организмов в том, что у первых отсутствуют полипептидные или олигопептидные гормоны, а в роли гормонов стали выступать менее сложные вещества, образующиеся, например, при модификации мономерных продуктов катаболизма белков (триптофан - ауксин, метионин – этилен) нуклеиновых кислот (пуриновые нуклеотиды - цитокинины) или липидов (жирные кислоты - жасмонат) [9, стр.559].
Большинство же физиологически активных олигомерных продуктов катаболизма биополимеров и оксигенированных производных полиеновых жирных кислот выполняет роль не гормонов, а внутриклеточных эндогенных регуляторов, не транспортируемых в другие органы и ткани. Они могут играть роль микровинта в корректировании метаболических процессов, в то время как грубая настройка осуществляется фитогормонами.
Еще один пример использования продуктов деградации биополимеров и липидов, выработавшийся в ходе эволюции, - это появление системы вторичных мессенджеров (инозитолтрифосфатов, диацилглицеролов, циклических нуклеотидов), функционирование которых показано в растениях и образование которых есть результат взаимодействия рецепторов плазмалеммы с гормонами и гидролиза мембранных фосфолипидов (в случае Са 2+-фосфоинозитольной системы), а также преобразования свободных нуклеотидов (синтез циклических нуклеотидов). Они, в свою очередь, вызывают фосфорилирование белков. Имеется мнение, что в результате фосфолирования белков протеинкиназами, активированными вторичными посредниками, повышается в целом активность ферментов катаболических реакций, фосфорилирование же ферментов анаболических реакций подавляет их активность [9, стр.557].
Особенно сильная активация катаболизма наблюдается при начинающемся стрессе. Известно, что ответная реакция растений на действие эколого-климатических (физических) и антропогенных (преимущественно химических), а также биогенных стрессоров включает несколько стадий:
1) тревоги,
2) адаптации,
3) истощения.
Одной из главных черт первой стадии и является резкое усиление процессов катаболизма липидов и биополимеров[8, стр.281].
Целесообразность усиления катаболизма определяется необходимостью повышения концентраций мономерных субстратов и ацетата для синтеза новых биополимеров и липидов, которые в большей степени соответствуют условиям стресса, а также необходимостью поддержания при этом энергетики клеток за счет сгорания продуктов катаболизма в процессе дыхания. Быстрое повышение концентрации субстратов для синтезов и обеспечение их макроэргическими соединениями и восстановителем создают условия для замещения одних распадающихся форм биополимеров и липидов другими[8, стр.289].
Еще один аспект катаболизма - включение сигнала тревоги - интенсивное образование олигомерных физиологически активных продуктов катаболизма полимерных молекул и оксигенированных производных липидов, в том числе тех фитогормонов, которые являются продуктами катаболизма. Эти сигналы тревоги приводят в действие сложную систему настройки клеток и организма в целом на экстремальные условия существования. Одним из компонентов этой системы настройки является усиление синтеза специфических (шоковых) соединений, способствующих повышению устойчивости клеток. В случае действия патогенов может наблюдаться, наоборот, сильное повышение чувствительности клеток вокруг места внедрения патогена и их гибель, но это повышает устойчивость организма, так как создает препятствия для дальнейшего распространения инфекции[8, стр.296].
Усиление деградации липидов и биополимеров - это лишь одна из многих неспецифических ответных реакций, отражающих структурно-функциональную перестройку растительного организма, попавшего в экстремальные условия.
Вполне естественно, что обнаруживаются и специфические черты ответа, зависящие от вида стрессора (в особенности от биогенного, от патогена), но по мере усиления меры его действия на первый план во все большей степени начинают выступать неспецифические изменения.
К числу наиболее значительных неспецифических изменений можно отнести следующие:
1. Фазность в развертывании во времени ответа на действие стрессора.
2. Усиление катаболизма липидов и биополимеров.
3. Изменение проницаемости мембран клеток для ионов.
4. Повышение в цитоплазме содержания ионов кальция.
5. Подкисление цитоплазмы.
6. Снижение общей интенсивности синтеза биополимеров и липидов.
7. Синтез стрессовых (шоковых) белков.
8. Интенсификация синтеза компонентов клеточных стенок - лигнина, суберина, кутина, каллозы, богатого оксипролином белка.
9. Накопление пролина, и связано это с его свойствами осморегулятора
10. Накопление органических полиаминов.
11. Повышение содержания абсцизовой и жасмоновой кислот.
12. Продукция этилена.
13. Торможение фотосинтеза.
14. Усиление дыхания.
15. Перераспределение углерода из СО2, усвоенного в процессе фотосинтеза, среди различных соединений - уменьшение включения метки в высокополимерные соединения (белки, крахмал) и сахарозу и усиление (чаще относительно - в процентах от усвоенного углерода) - в аланин, малат, аспартат. Положение о неспецифических изменениях в углеродном метаболизме, вызванных действием различных стрессоров, и о непосредственных причинах этих изменений было впервые выдвинуто автором более четверти века тому назад [1, стр.3].
Похожие работы
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... городская конвекция воздуха от более тёплого центра, где «остров тепла», к более холодным окраинам, и от жилых кварталов к паркам и водоёмам создаёт постоянные ветры типа городских бризов, то проблема иссечения растений частицами очень существенная. В Европе, где степень урбанизации много выше нашей (урбанизированные районы сливаются, образуя мегалополисы с населением 20-50 млн. человек, и уже ...
... и утрачивается к началу полового созревания. 36. Учение Павлова о типах ВНД. Общин типы ВНД. Пластичность ВНД. Учет типов ВНД в индивидуальном обучении и воспитании Среди вопросов физиологии ВНД человека особое значение для совершенствования учебно-воспитательного процесса и разработки естественнонаучных теорий воспитания и обучения имеет учение о типах ВНД. Это связано с тем, что именно ...
... агент в месте его внедрении. На МАС, как правило, накладывается ГАС, образованный развивающимися общими явлениями. Наоборот, ГАС вторично влияет на МАС с помощью нейрогуморальных механизмов (например, антивоспалительных гормонов). Шок (англ. shock – удар) – патологический процесс, возникающий при действии на организм сверхсильных патогенных раздражителей и характеризующийся фазным нарушением ...
0 комментариев