Навигация
Повышение содержания свободных радикалов
39242
знака
0
таблиц
3
изображения
16. Повышение содержания свободных радикалов.
17. Образование в ответ на действие биострессоров (бактерии, грибов, вирусов, насекомых) элиситоров и фитоалексинов фенилпропаноидной и терпеноидной природы, разнообразных патогениндуцированных белков, хитиназ, β-1, 3-глюканаз, ингибиторов протеиназ[1, стр.4].
Неблагоприятные условия существования - климатические, биогенные, антропогенные- действуют на обмен веществ и ультраструктуру клеток в том же направлении, что и старение. Более того, они ускоряют его. Можно считать, что описания особенностей процессов быстрого старения у таких обычно используемых для этого объектов, как лепестки цветов в значительной степени относятся и к растениям, подвергшимся действию неблагоприятных факторов. Это в первую очередь касается отсеченных лепестков, часто используемых в качестве модели для изучения старения. Дело в том, что отделение какого-либо органа или вычленение ткани растения само по себе вызывает у них состояние стресса. Это продемонстрировано на отделенных листьях и особенно полно на отсеченных корешках проростков.
Например, в некоторых работах показано, что фазы стресса (реакция тревоги и реакция адаптации) реализуются в процессе адаптивного старения отсеченных корней [2, стр.120]. Отсечение корня от проростка вызывает в начальный период (1-2 ч после отсечения) ряд изменений в метаболизме, которые можно отнести к реакции тревоги или предадаптации. Это - падение мембранного потенциала, усиление генерации активных форм кислорода, повышение выхода из клеток ионов калия (вероятно, по калиевым каналам), увеличение содержания свободных жирных кислот, повышение степени ненасыщенности жирнокислотных остатков мембранных фосфолипидов, активация митохондриального дыхания.
Все эти изменения рассматриваются как основа последующих восстановительных процессов, которые можно отнести к стадии адаптации. Во время стадии адаптации происходят восстановление мембранного потенциала, снижение генерации активных форм кислорода, поглощение ионов калия (возможно, вследствие активации протонных насосов). Одновременно повышается степень насыщенности жирнокислотных остатков мембранных фосфолипидов, что является основой снижения степени проводимости мембран для ионов [3, стр.57].
Старение организмов приводит к постепенному уменьшению величины отношения анаболизм/катаболизм. В различных органах растений степень усиления катаболизма с увеличением общего возраста организма может отличаться в зависимости от собственного возраста, например, у метамерных органов (листьев). Усиление катаболизма старых листьев способствует «перекачке» образующихся мономеров (и продуктов их превращения в легко транспортируемые формы) и реутилизации органических соединений в более молодых или репродуктивных органах. Среди неспецифических ультраструктурных изменений, вызванных стрессом и старением, можно отметить изменение тонкой гранулярной структуры ядра, уменьшение числа полисом и диктиосом, набухание митохондрий и хлоропластов, уменьшение в хлоропластах числа тилакоидов, перестройку цитоскелета [3, стр.157].
В изучении процессов деградации у растений важную роль играет выбор объекта исследований. Например, сильная интенсификация катаболизма биополимеров и липидов наблюдается у прорастающих семян; в органах растений с быстрым темпом старения (например, у лепестков некоторых видов растений); у затемненных листьев; в участках корней, заканчивающих рост растяжением [4, стр.197]; в тканях и клетках, разрушающихся в результате механического повреждения, подвергающихся действию бактерий, грибов, вирусов, эколого - климатических или антропогенных стрессоров. Во многих случаях используются изолированные протопласты, культуры клеток или каллусных тканей растений.
Анализ событий, совершающихся в клетках с момента начала действия того или иного стрессора, заставляет считать, что имеется небольшое число наиболее лабильных (чувствительных) компонентов клеточных структур и звеньев метаболизма, которые выполняют роль пускового механизма, вызывающего последующие глобальные нарушения, проявляющиеся в виде неспецифических изменений в обмене веществ.
По всей вероятности, функцию спускового крючка, на которых "нажимают" разнообразные стрессоры и который приводит в действие сложный механизм ответа, в большинстве случаев выполняют мембранные системы.
Мембраны - это "ахиллесова пята" клеток, в первую очередь повреждающаяся под влиянием экстремальных факторов. Совершенно очевидно, что во многих случаях, например, при изменении температуры, роль сенсорного компонента мембран играют липиды, от состава и соотношения которых зависит степень вязкости мембран, определяющая структуру и функции мембранных белков.
Изменение состояния мембран приводит к включению процесса деградации липидов, от которого зависит реализация программы формирования стресса растительных клеток[4, стр.197].
3. БИОГЕННЫЙ СТРЕСС И КАТАБОЛИЗМ
В предыдущем разделе уже затрагивался вопрос о влиянии патогенов на образование углеводных элиситоров и белковый обмен. В последние годы появляется все больше информации о метаболическом взаимодействии патогенных организмов - бактерий и грибов - и тканей различных органов растения-хозяина. Первый этап этого взаимодействия - конформационное узнавание поверхностей органов и клеток патогена и хозяина. Второй - экскреция клетками патогенных бактерий и грибов ферментов, гидролизующих биополимеры и липиды покровных тканей растения-хозяина, разрыхляющих их и обеспечивающих более интенсивное проникновение патогена в ткани хозяина и обильное питание, необходимое для развития патогена. Третий - образование в ходе деградации биополимеров и липидов различных физиологически активных промежуточных продуктов - элиситоров, выполняющих роль сигнальных веществ - стимуляторов ответной реакции клеток хозяина. Элиситоры непосредственно или с помощью посредников влияют на генетический аппарат клеток хозяина, вызывая (четвертый этап) синтез веществ, способствующих или повышению устойчивости к патогену, или вызывающих сверхчувствительность и гибель клеток, но тем самым создающих механический барьер, препятствующий распространению инфекции по тканям растения (рис. 2). Промежуточные продукты катаболизма клеток хозяина могут выступать и в роли эффекторов метаболизма и развития патогена [5, стр.44].
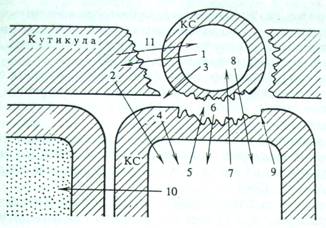
Рис. 2. Схема взаимодействия клетки патогена с растением- хозяином
1 — кутиназа; 2 — продукты деградации компонентов кутикулы (возможно, обладающие сигнальными свойствами); 3 — β-глюканаза и другие гликозилазы, экскретируемые патогеном; 4 — элиситоры — фрагменты клеточной стенки (КС) хозяина; 5 — хитиназы и другие гликозилазы, действующие разрушающе на КС патогена; 6 — элиситоры — фрагменты КС патогена; 7 — фитоалексины — ингибиторы протеиназ, кутиназ, гликозилиз и других ферментов патогена; 8 — токсические вещества патогена; 9 — укрепление КС хозяина за счет активации пероксидаз и усиления синтеза лигнина. Отложение оксипролиновых белков, лектинов; 10 — вещества — индукторы сверхчувствительности и некроза соседних клеток; 11 — продукты деградации кутина, действующие на клетку патогена
Наружным покровом растений является кутикула, состоящая главным образом из гетерополимера кутина, погруженного в воск. Обнаружено более 20 мономеров, из которых состоит кутин: различной длины насыщенные и ненасыщенные жирные кислоты и спирты, в том числе гидроксилированные и эпоксидированные, дикарбоксиловые кислоты и т.д. В кутине большинство первичных спиртовых групп участвует в образовании эфирных связей, так же как часть вторичных спиртовых групп, обеспечивающих сшивки между цепями и точки ветвления в полимере. Другой "барьерный" полимер - суберин, состоит, по-видимому, из фенольных и алифатических доменов, первые из которых близки по своему составу к лигнину, а вторые - к кутину. Отличия алифатического домена в том, что свободные жирные кислоты являются главным компонентом субериновых восков, в то время как в кутине их очень мало. Кроме того, в суберине присутствуют главным образом С22и С24 жирные спирты, в то время как в кутине – С26 и С 28 [5, стр.44].
Оказалось, что многие патогенные грибы могут выделять ферменты, гидролизующие кутин и суберин. При гидролизе лигниноподобной фракции суберина образовывались кумаровая и феруловая кислоты, причем большая часть фракции оставалась негидролизованной. Продуктами кутиназной реакции были различные оксигенированные жирные кислоты и спирты [5, стр.45].
По всей вероятности, в спорах грибов кутиназа содержится в очень небольших количествах, и при контакте с кутикулой растений гидролизу подвергается лишь малая часть кутина. Однако образующиеся активные сигнальные молекулы - 10,16-дигидрокси-С16- и 9,10,18-тригидрокси-С18-кислоты - транспортируются в прорастающую спору и индуцируют образование больших количеств дополнительной кутиназы, начинающей интенсивное разложение кутина и облегчающей инфицирование растения. Было обнаружено, что лаг-период появления кутиназной м-РНК после начала действия ди- и триоксикислот составляет всего 15 мин, а появления кутиназы - в два раза больший [5, стр.45]. Ингибирование кутиназы с помощью химических препаратов или антител предотвращало инфекцию.
Одной из важных задач исследователей молекулярных взаимодействий между патогеном и тканями растения-хозяина были поиски сигнальных веществ, индуцирующих защитную реакцию инфицированных клеток и системную - в удаленных от места инфекции местах.
Вполне вероятно, что продукты деградации кутина (оксигенированные жирные кислоты и спирты) могут выступать в роли не только индукторов образования кутиназы у патогена, но и элиситоров синтеза защитных веществ в клетках растения-хозяина (см. рис.2).
К числу высокоэффективных элиситоров защитного ответа инфицированных растений относятся олигомерные продукты деградации полисахаридов клеточных стенок хозяина или патогена[5, стр.45].
Гидролитические ферменты (в том числе гликопротеины) патогенных грибов активируют защитную реакцию хозяина, освобождая биологически активные углеводы из клеточных стенок хозяина или патогена.
Было найдено, что суспензионные клетки табака вырабатывали фитоалексины (капсидиол и др.) в ответ на обработку целлюлазой, что свидетельствует об элиситорных свойствах линейных β-1,4-глюкановых фрагментов целлюлозы. В этой связи представляют интерес данные об активирующем действии дисахарида целлобиозы на синтез целлюлозы из 14С-глюкозы или из меченой УДФГ в клетках волосков семян хлопчатника. Так как в клетках не содержится эндогенной целлобиозы, то, по-видимому, экзогенная целлобиоза служит миметиком олигосахаридов более высокой степени полимеризации[9, стр.562].
В формировании ответной реакции растений на патогены принимают участие также полиеновые жирные кислоты.
Оказалось, что элиситорным эффектом обладает не белковая часть липопротеинов, а их липидная часть, представляющая собой не свойственные для высших растений арахидоновую (эйкозатетраеновую) и эйкозопентаеновую кислоты. Они вызывали образование фитоалексинов, некротизацию тканей и системную устойчивость растений к различным патогенам. Продукты липоксигеназного превращения в тканях растений упомянутых выше С20 жирных кислот (гидроперокси-, гидрокси-, оксо-, циклические производные, лейкотриены), образующиеся с помощью имеющегося в клетках хозяина ферментного липоксигеназного комплекса (субстратами которого могут быть как C18, так и С20 полиеновые жирные кислоты), оказывали сильнейшее влияние на защитную реакцию растений. Это объясняется, по-видимому, тем, что в неинфицированных растениях нет оксигенированных производных 20-углеродных жирных кислот, и их появление в результате инфицирования приводит к драматическим результатам, например к гибели клеток и образованию некрозов, что создает барьер для распространения инфекции. Имеются данные, что индуцирование патогеном липоксигеназной активности приводило к формированию ответной реакции растения и в том случае, когда элиситор не содержал С20 жирных кислот и субстратом липоксигеназной активности могли быть только собственные полиеновые жирные кислоты, а продуктами - октадеканоиды, а не эйкозаноиды. В связи с этим представляют большой интерес сведения о том, что жасмонат может индуцировать синтез ингибиторов протеиназ[10, стр.562]. Знаменательно, что глюкан и Са2+ усиливали влияние арахидоната и эйкозапентаеноата. Так как ЭГТА (специфический лиганд Са2+) ингибировал синтез фито-алексинов, то можно сделать предположение, что ионы кальция играют важную роль в регуляции осуществления защитной функции растений[5, стр.44].
Не исключено, что сигнальными веществами являются и продукты деградации белков клеточных стенок, богатых оксипролиновыми остатками и содержащих олигогликозильные ответвления.
Под влиянием воздействия многообразных элиситоров в инфицированных растениях начинают вырабатываться защитные вещества, повышающие устойчивость клеток к инфекции. К ним относятся в первую очередь фитоалексины - растительные антибиотики, представляющие собой соединения фенилпропаноидного и терпеноидного характера. Например, у картофеля под влиянием Phytophtora infestans образуются ришитин и любимин[2, стр.123].
Элиситоры индуцируют образование большого количества белков, как правило, не характерных для неинфицированных тканей.
1. Патогениндуцированные белки, представляющие собой группу щелочных и кислых белков с относительно небольшой молекулярной массой (10-20 кДа), функции которых в большинстве случаев не выяснены.
2. Хитиназы и β -глюканазы, накапливающиеся в вакуоле и клеточных стенках. Интересно, что b -глюканаза "созревает" в две стадии. Первая заключается в удалении с N-конца олигопептида (насчитывающего 21 остаток аминокислот) и в N-гликозилировании С-конца белка, вторая - в отщеплении от образовавшегося промежуточного белка олигопептида с 22 аминокислотными остатками, включая углеводную ветвь, присоединившуюся на предыдущей стадии.
3. Ингибиторы протеиназ, вырабатываемые как в результате механического повреждения тканей (например, листогрызущими насекомыми), так и в результате инфицирования патогенами. Их синтез вызван фрагментами пектиновых веществ клеточных стенок. Интересно, что индукция ингибиторов протеиназ сопровождалась фосфорилированием белков плазмалеммы клеток хозяина.
Похожие работы
... , часть накопленных в процессе фотосинтеза углеводов теряется. Однако все попытки искусственно ингибировать фотодыхание приводили к общему снижению интенсивности фотосинтеза. На современном этапе развития физиологии растений принято считать, что основное значение фотодыхания заключается в его защитной роли. Сбрасывая таким образом избыточную энергию, растение избегает разрушения фотосистем, ...
... городская конвекция воздуха от более тёплого центра, где «остров тепла», к более холодным окраинам, и от жилых кварталов к паркам и водоёмам создаёт постоянные ветры типа городских бризов, то проблема иссечения растений частицами очень существенная. В Европе, где степень урбанизации много выше нашей (урбанизированные районы сливаются, образуя мегалополисы с населением 20-50 млн. человек, и уже ...
... и утрачивается к началу полового созревания. 36. Учение Павлова о типах ВНД. Общин типы ВНД. Пластичность ВНД. Учет типов ВНД в индивидуальном обучении и воспитании Среди вопросов физиологии ВНД человека особое значение для совершенствования учебно-воспитательного процесса и разработки естественнонаучных теорий воспитания и обучения имеет учение о типах ВНД. Это связано с тем, что именно ...
... агент в месте его внедрении. На МАС, как правило, накладывается ГАС, образованный развивающимися общими явлениями. Наоборот, ГАС вторично влияет на МАС с помощью нейрогуморальных механизмов (например, антивоспалительных гормонов). Шок (англ. shock – удар) – патологический процесс, возникающий при действии на организм сверхсильных патогенных раздражителей и характеризующийся фазным нарушением ...
0 комментариев