Навигация
Асимметрия мембран
Введение
Все биологические мембраны асимметричны, и легко понять почему: ведь каждая из них имеет две поверхности, омываемые разными средами. В качестве примера можно привести плазматическую мембрану, обращенную одной стороной в цитоплазму, а другой — во внеклеточное пространство. Именно трансмембранная асимметрия, дифференцирующая две половинки бислоя, обусловливает чувствительность мембраны к изменениям среды по обе ее стороны. Очевидно, что асимметрия мембранных белков зависит от способа, каким тот или иной белок был внедрен в мембрану. Скорость «флип-флопа» белков в бислое пренебрежимо мала. Мембранные липиды тоже расположены асимметрично. Наиболее убедительно это было показано для эритроцитов. Как возникает липидная асимметрия и как она поддерживается — в настоящее время во многом неясно. В одних случаях важную роль играют физические факторы, в частности кривизна мембраны, в других определяющий вклад вносят взаимодействия с цитоскелетом или участие АТР-зависимых ферментоподобных «флипаз».
Ясно, что помимо трансмембранной асимметрии мембранам присуща и латеральная негомогенность. Поверхностная мембрана многих эукариотических клеток сильно поляризована и имеет четко выраженные макроскопические домены. В качестве примера можно привести базолатеральную и апикальную области плазматической мембраны поляризованных эпителиальных клеток. Эти домены выполняют различные функции, имеют неодинаковый состав и физически разнесены по поверхности клетки. Мембраны тила-коидов в хлоропластах также имеют доменную структуру; в частности, у них есть плотно прилегающие друг к другу мембранные участки и несоприкасающиеся области, содержащие различные элементы системы электронного транспорта. Эти достаточно протяженные латеральные домены могут существовать за счет специфических белок-белковых взаимодействий между мембранами или могут быть обусловлены наличием в самой мембране специальных структур, взаимодействием с компонентами цитоскелета или агрегацией белков в плоскости мембраны. Детали этих взаимодействий пока неизвестны.
Что касается возможности существования малых липидных доменов в мембране, т. е. отдельных участков, в пределах которых бис-лой имеет специфические физические свойства и состав, то говорить об этом можно с меньшей определенностью. В модельных системах при различных условиях действительно наблюдается латеральное разделение фаз, и заманчиво было бы предположить, что подобное разделение существует и в биологических мембранах при физиологических условиях. Возможно, это не-бислойные элементы мембраны или какие-то переходные структуры.
Мы рассмотрим асимметричное распределение мембранных белков и липидов. В целом всю эту область можно назвать мембранной топографией. Сюда же относится и описание цитоскелета, поскольку он, по-видимому, играет важную роль в организации белков и, возможно, липидов в плазматической мембране эукариот.
1. Топография мембранных белков
Мембранные белки встроены в бислой асимметрично, и эта асимметрия регулируется кинетическими факторами. Энергия активации для такой переориентации белка в мембране, когда полярные и заряженные остатки, обычно находящиеся на поверхности, хотя бы временно оказываются в гидрофобной области бислоя, очень велика. И хотя известно, что некоторые мембранные белки диффундируют вдоль бислоя и/или вращаются вокруг оси, перпендикулярной его поверхности, нет никаких данных о том, что какое-либо спонтанное перемещение может привести к изменению транс-мембранной ориентации белка по отношению к двум сторонам бислоя.
В этом разделе представлен обзор основных экспериментальных подходов к определению трансмембранной ориентации белков в би-слое. Есть много способов, позволяющих предсказать локализацию спиралей, пронизывающих мембрану, исходя из первичной последовательности данного белка. В одних случаях эти предсказания являются достаточно четкими и согласуются с результатами экспериментальных исследований, в других однозначный ответ получить не удается. Ни одна модель, построенная с помощью компьютера, не является окончательной, она дает лишь исходную точку для анализа. Совершенно ясно, что любое детальное исследование интегрального мембранного белка должно включать экспериментальное изучение его топографии. Сделать это бывает непросто, и часто для построения адекватной модели приходится применять несколько подходов, поскольку ни один метод не застрахован от ошибок. Детальному рассмотрению отдельных экспериментальных подходов посвящены обзоры.
1.1 Методология протеолиз
Использование протеаз для определения топографии белков в ряде случаев оказалось исключительно плодотворным. Ориентированный мембранный препарат, содержащий изучаемый белок, обрабатывают протеолитическим ферментом и по местам расщепления устанавливают те участки полипептида, которые находятся с наружной стороны мембраны. Ключевым моментом является приготовление мембран с однозначной топологической ориентацией; только в этом случае возможна адекватная интерпретация результатов фрагментации изучаемого белка. Некоторые мембраны можно изучать без предварительного выделения. Однако в большинстве случаев необходима тщательная работа по получению топологически ориентированного препарата с вывернутой мембраной или с мембраной, имеющей нативную ориентацию. Такие препараты были получены для мембран эритроцитов, плазматических мембран некоторых эукариотических клеток, внутренних митохондриальных мембран, мембран саркоплазматического рети-кулума и некоторых бактериальных мембран. В отдельных случаях удается исследовать локализацию конкретного белка, специально встроенного в ориентированную мембрану реконструированных протеолипосом. В биологических мембранах помимо изучаемого белка чаще всего находятся и другие белки. Если изучаемый белок преобладает, то продукты протеолиза можно выделить и охарактеризовать. Если же он является минорным белковым компонентом, то для идентификации фрагментов приходится использовать косвенные методы, связанные с применением специфических антител или химических реагентов с последующим электрофорезом в ПААГ в присутствии ДСН. Важно убедиться в том, что те участки белка, которые недоступны для расщепления, не будут подвергаться гидролизу из-за повреждения мембран при первоначальном протеолизе. Необходимо также быстро и обратимо ингибировать протеа-зы, поскольку многие из этих ферментов остаются активными даже после добавления ДСН перед электрофорезом. Следует иметь в виду, что отсутствие расщепления еще ничего не означает, поскольку возможные места расщепления на наружной поверхности могут оказаться недоступными для протеазы из-за особенностей третичной структуры мембранного белка.
Как и в случае многих растворимых белков, протеолиз не обязательно сопровождается существенными изменениями в третичной или четвертичной структуре мембранного белка in situ, хотя его биологическая функция может быть полностью утрачена. В тех случаях, когда при расщеплении по одному или двум местам активность исчезает, с помощью протеолиза можно идентифицировать и даже выделить функционально важные домены белка.
В качестве примеров, иллюстрирующих использование протеоли-тических ферментов, можно привести белок полосы 3 эритроцитов, бактериородопсин, лактозопермеазу Е. coli и субъединицу IV цитохром с-оксидазы.
Иммунологические методы
Очень ценным инструментом для определения топографии мембранных белков являются специфичные антитела. В этом случае исследуют связывание антител с белками только в ориентированных мембранных препаратах типа мембранных везикул Е. coli. Анализ можно сделать количественным, если использовать соответствующие иммунологические методы. Места связывания можно локализовать с помощью электронной микроскопии, пометив антитела коллоидным золотом или используя золото, связанное со специфичным комплексом антиген—антитело на поверхности мембраны. Ясно, что в процессе приготовления мембранного препарата не должны разрушаться нативные топографические структуры. Чем точнее данные о местах связывания антител, тем информативнее будут эти эксперименты. Если исследовать ориентированные мембраны с нормальной или вывернутой ориентацией, то с помощью поли-клональных антител против определенного очищенного полипептидного фрагмента можно определить, имеет ли изучаемый белок участки, экспонированные на какой-либо одной стороне мембраны. Однако такие эксперименты не позволяют определить, какая часть белка экспонирована. Более детальную информацию можно получить с помощью двух подходов. Один состоит в использовании мо-ноклональных антител, а второй предусматривает применение поли-клональных антител против пептидов, соответствующих определенным областям белка.
Наиболее полезны те из них, которые связываются как с белком в мембране, так и с денатурированными фрагментами белка, что позволяет изучать белки после их разделения с помощью электрофореза в ПААГ в присутствии ДСН. Это особенно важно для локализации места связывания антител на полипептиде. В принципе эпитоп можно локализовать с точностью до нескольких аминокислотных остатков. В качестве примеров успешного использования моноклональных антител можно привести родопсин, бактериородопсин, ацетилхолиновый рецептор и белок LamB — рецептор фага X из наружной мембраны Е. coli.
Использование антител против синтетических пептидов, соответствующих отдельным белковым фрагментам, позволяет точно выяснить, доступна ли эта область белка для связывания антитела. Если антитела, специфичные к определенной аминокислотной последовательности, связываются с нативной формой белка в мембране, то исследователь получает мощный инструмент для определения топографии белка. Этим путем можно проверять отдельные топографические модели. Примерами успешного использования данного подхода являются работы по лактозопермеазе Е. coli, микросомному цитохрому Р450 и ацетилхолиновому рецептору. К сожалению, нет гарантий, что антипептндные антитела вообще будут связываться с белком. Потенциальный центр связывания в нативном белке может находиться в такой конформации или так быть упрятанным внутри белковой глобулы, что не будет узнан антителом или доступен для него. Действительно, в некоторых случаях антипептидные антитела не связываются с белком даже после его денатурации в ДСН.
Весьма остроумным является подход к определению топографии полипептида, основанный на методах молекулярной генетики. Он состоит во введении чужеродного эпитопа в аминокислотную последовательность мембранного белка с последующим использованием антител, специфичных к этому эпитопу.
Химическая модификация
Этот подход широко использовался для локализации белков или их отдельных участков на поверхности мембраны или в ее гидрофобной области. Белок вступает в реакцию с реагентом, который может действовать лишь на одной стороне ориентированной мембраны или в ее гидрофобной сердцевине. Реагент, предназначенный для идентификации белка на поверхности мембраны, должен быть сильно полярным; он не должен адсорбироваться на мембране или накапливаться внутри ее. Для локализации же той части белка, которая находится в контакте с углеводородными хвостами липидных молекул, следует использовать очень неполярные реагенты, которые концентрируются в гидрофобной области мембраны. Артефакты, возникающие при этом подходе, чаще всего бывают связаны с тем, что реагент модифицирует белок не только в том реакционном пространстве, для которого он был предназначен. Например, реагент, направленный на модификацию поверхности мембраны, может иметь достаточно неполярный характер, чтобы проникать через мембрану и получать доступ к белкам внутреннего компартмента. Химическая модификация может также привести к повреждению мембраны и сделать ее проницаемой для того реагента, который, как ожидается, должен быть непроникающим. В табл. 1 перечислены некоторые из химических реагентов, используемые для изучения топографии мембранных белков.
Модификацию белков на поверхности мембраны часто проводят с помощью фермента лактопероксидазы, которая катализирует иодирование доступных остатков тирозина или гистидина. Поскольку реакция между I" и Н2О2 происходит в активном центре лактопероксидазы, последняя должна быть в высшей степени «векторной», т.е. локализованной только на одной стороне мембраны. Однако при определенных условиях в ходе реакции может образовываться Ь, который, проникая через мембрану, способен иодировать аминокислотные остатки на внутренней стороне мембраны. Это показывает, насколько важен строгий контроль условий протекания реакций для исключения артефактов.
Для поверхностной модификации часто используются и другие реагенты — соли диазония, которые реагируют с боковыми цепями остатков лизина, цистеина, тироксина и гистидина, а также фотоак-тивируемые реагенты, например NAP-таурин.
В табл. 1 перечислен также ряд неполярных фотоактивируемых реагентов, которые используются для избирательной модификации аминокислотных остатков белка, контактирующих с гидрофобной областью бислоя. Эти реагенты обычно добавляют к мембранному препарату, дают им возможность накопиться в бислое и затем подвергают их фотоактивации. Преимущество образующихся при этом нитренов и карбенов состоит в том, что их реакции намного менее специфичны по сравнению с реакциями других активных частиц, так что ковалентная модификация не ограничивается боковыми цепями каких-либо определенных аминокислот. Правда, нитрены проявляют избирательность по отношению к нуклеофилам. Использование карбенов предпочтительнее, поскольку они вступают в реакцию с более высоким выходом. Для получения информации о вторичной структуре трансмембранных участков полипептидной цепи путем определения тех остатков, которые контактируют с липидами, использовали реагент ТИД. Например, спираль С бак-
Таблица 1. Некоторые реагенты, применяемые для изучения топографии мембранных белков
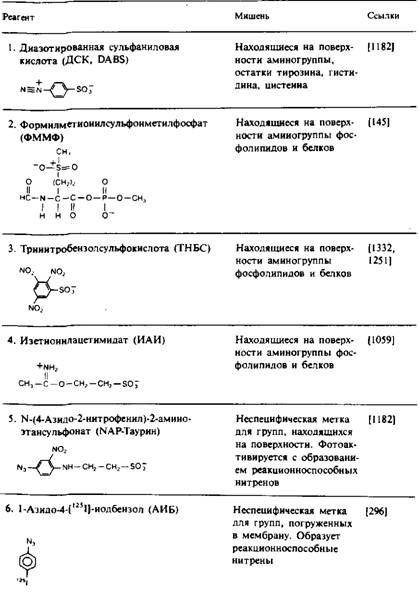

териородопсина включает метку лишь с одной стороны; это согласуется с представлением о том, что данная спираль является трансмембранной, причем ее неполярная область обращена в липидную фазу.
Локализация специфических центров
Ценную информацию иногда можно получить, определяя положение специфических центров в ряде белков. Например, места присоединения сахарных остатков в гликопротеинах плазматической мембраны всегда находятся на ее наружной поверхности, так что выявление их в полипептидной цепи имеет и топологическую ценность. Столь же информативными могут быть и данные о локализации сайтов модификации белка, например сайтов фосфорилирования, если локализация модифицирующего фермента известна. Так, в белках плазматической мембраны фосфорилированные аминокислотные остатки находятся на цитоплазматической стороне. Аналогичным образом может оказаться полезной локализация специфических мест связывания. Например, были идентифицированы места связывания бактериофага с экспонированными на поверхности клетки участками белков ОтрА и LamB наружной мембраны Е. coli; для этого использовались мутантные белки.
Генетические подходы
Возможность генетической модификации мембранных белков привела к созданию новых подходов к их топографическому анализу. Такие методы, по всей вероятности, будут применяться все шире, однако пока число удачных примеров не столь велико, чтобы судить об их надежности и универсальности. Так, в белок ОтрА методами генной инженерии были встроены в определенные места короткие пептиды, затем был проведен протеолиз и по его результатам определено, были ли участки, содержащие включенный пептид, экспонированы на поверхности клетки. Анализ мутантных вариантов позволил идентифицировать также места связывания фага и соответствующие эпитопы в белках, экспонированных на наружной стороне мембраны.
Ценные данные о топографии белков цитоплазматической мембраны Е. coli были получены и с помощью метода гибридизации белков. Гибридные белки можно сконструировать так, что их N-концевой участок будет представлен мембранным белком, а на С-конце будет находиться каталитический центр щелочной фосфатазы. Щелочная фосфатаза обычно локализуется в периплазматическом пространстве Е. coli, куда она транспортируется и где проявляет свою ферментативную активность. Синтез многих мембранных белков, по-видимому, осуществляется линейным образом, начиная с N-конца. Поэтому, если место сочленения, где начинается последовательность щелочной фосфатазы, локализовано на периплаз-матической стороне мембраны, то гибридный белок будет транспортироваться в периплазму и проявлять ферментативную активность. Если же место сочленения находится на цитоплазматической стороне, то щелочная фосфатаза в гибридном белке останется внутри клетки и будет проявлять низкую ферментативную активность. Следовательно, те места сочленения в гибридных белках, которые приводят к высокой активности щелочной фосфатазы, будут соответствовать наружным доменам мембранного белка.
Похожие работы
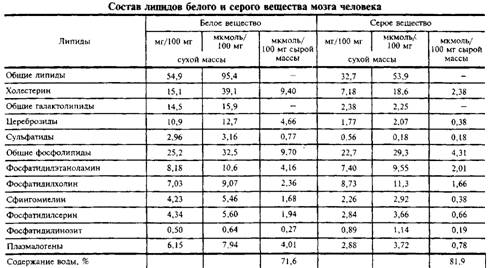
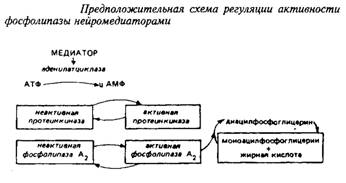
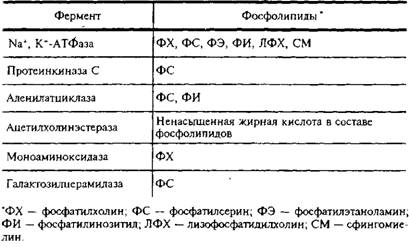
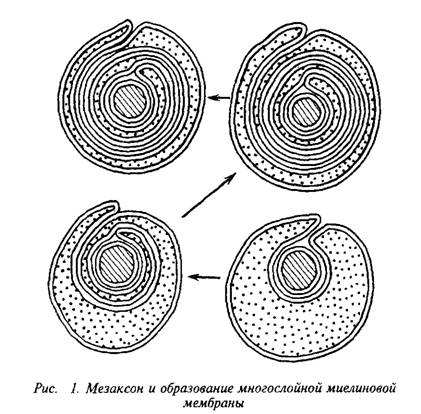
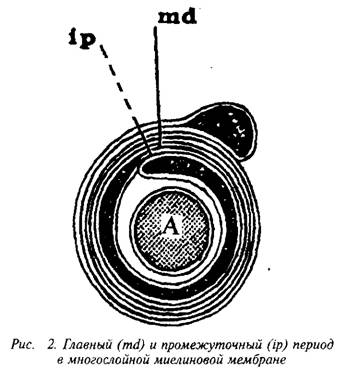
... и инозитолтрифосфат подвергаются химическим превращениям, требующим АТФ и ЦТФ и приводящим к восстановлению три-фосфоинозитида. Таким образом, цикл замыкается и уровень полифосфоинозитидов в мембране восстанавливается. 7. МИЕЛИН В ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЕ Мозг человека содержит 120 г миелина, что составляет одну треть его сухой массы. Миелин – уникальное образование, организация которого ...
... является курсом, для изучения которого необходимо наличие знаний об основных принципах организации биологических молекул, строении и механизмах действия ферментов. Дисциплина биохимия мембран относится к дисциплинам специализации федерального компонента. 5. Распределение времени, отведенного на изучение дисциплины по учебному плану Форма учебной работы Форма обучения Очная По ...
... молекул детергента приходится на одну мицеллу. В мембранных исследованиях используют довольно ограниченный круг детергентов. В табл. 1 представлены те из них, которые чаще всего применяются для солюбилизации и реконструкции мембран. Для этих детергентов характерны довольно высокие значения ККМ (10- 4-10- 2 М) и то, что они относятся к разряду так называемых мягких детергентов, то есть таких, ...
... широко известным из которых является электрофорез. Все вышеизложенное позволяет отнести тему нашей работы по исследованию механизма проницаемости плацентарных мембран по анионам антибиотиков в малоамплитудных физических полях к новому актуальному научному направлению – электрохимической кинетике. Цель работы На основании теоретических и экспериментальных исследований определить особенности ...
0 комментариев