Навигация
Структура животного населения почвы и факторы его разнообразия
71949
знаков
3
таблицы
6
изображений
1.2 Структура животного населения почвы и факторы его разнообразия
В наземных экосистемах во всех районах Земли обитателями почвы являются 50—99% всех видов животных и на их долю приходится 60—90% наземной зоомассы. Число особей на единицу площади у некоторых групп достигает фантастических величин (до 1 млн. клещей, ногохвосток на 1 кв. м в лесных и луговых почвах). Причины высокого обилия различных представителей почвенного населения изложены в книгах М. С. Гилярова.
Почва представляет очень сложную, многокомпонентную среду. Это трехфазная и полидисперсная система, в которой промежутки между твердыми частицами и их агрегатами заполнены воздухом и водой с растворенными в ней солями. Полидисперсность почвы, или ее гранулометрия, выражается в содержании механических элементов разного размера, от отдельных ионов почвенного раствора до каменистых включений. Разное соотношение фаз создает гамму условий и поэтому для разных размерных групп организмов почва представляет неодинаковую среду, что является одним из факторов высокого разнообразия ее биоты.
В почвенной зоологии принято выделять размерные группы животных, различающихся способами использования среды обитания:
- нанофауна (размеры от микрон до долей мм) – микроскопические объекты, которые могут наблюдаться только с помощью инструментальных методов: простейшие (корненожки, жгутиконосцы, инфузории), мелкие нематоды и коловратки, тихоходки;
- микрофауна (доли мм – доли см) — более крупные немикроскопические организмы: клещи, нематоды, энхитреиды, пауки, коллемболы (ногохвостки), протуры, симфилы, мелкие жуки;
- мезофауна (доли см – несколько см) — крупные беспозвоночные, хорошо различаемые невооруженным глазом, легко учитываемые в полевых условиях при ручной разборке проб почвы: кольчатые и плоские черви, многоножки, пауки, мокрицы, брюхоногие моллюски, насекомые на разных стадиях развития;
- макрофауна (мегафауна) — почвенные позвоночные и крупные формы беспозвоночных.
Установлена обратная зависимость между размерами животных и уровнем их численности (М. С. Гиляров).
Разные размерные группы животных неодинаково используют почву как среду обитания (М. С. Гиляров). Для микроскопических организмов средой обитания оказывается не вся почва, а система микрокапель, капилляров, гравитационной воды, скопления влаги на твердых частицах и между частицами. Когда в почве имеется капиллярная и гравитационная вода, простейшие и коловратки плавают в ней. Животные сохраняют жизнеспособность даже в пленочной воде, находясь в неподвижном состоянии, но не прекращая питания микроорганизмами, детритом, оказавшимися в тех же водяных пленках. Обитатели пленочной воды почв входят также в состав фауны грунтов пресноводных водоемов (Д. А. Криволуцкий). Хотя в почве распределение влаги неравномерное, «прерывистое», но общая поверхность различных форм воды столь велика, что ее можно считать специфическим водоемом, а микроскопических обитателей — физиологически водной экологической группой. Существенное значение для этих организмов имеют особенности почвенной влаги: реакция (рН), химический и газовый состав, наличие почвенных коллоидов, состав водорастворимых солей, особенности органического вещества и порового пространства.
Для мелких членистоногих, называемых микроартроподами, среда обитания — это система ходов и полостей между частицами почвы и их агрегатами, почвенные трещины, ходы более крупных животных и корней, заполненные влажным воздухом. Условия жизни в почве этой группы животных М.С. Гиляров сравнивает с обитанием в насыщенных влагой пещерах. Передвижение обитателей таких пустот не отличается от передвижения по поверхности твердого субстрата. Они могут переживать периоды затопления почвы в отдельных пузырьках воздуха. Все сказанное позволяет считать их физиологически строго наземной экологической группой. Наибольшее значение для этой категории имеют степень порозности и влажности среды, характер распределения органических остатков и гумуса и температурный режим.
Для крупных животных — беспозвоночных и позвоночных — почва представляет среду обитания в целом. Она выступает как рыхлый или плотный и даже твердый субстрат. Движение в плотных слоях почвы встречает большое сопротивление. Передвижение возможно либо по естественным скважинам для животных с тонким змеевидным телом, либо раздвигая частицы почвы, либо копая и прогрызая ходы. При этом животные всегда испытывают механическое воздействие ее твердой фазы и химические воздействия почвенного раствора через покровы, особенно в периоды переувлажнения или затопления почвы. Для крупных почвенных животных имеет значение вся совокупность свойств почвы как единого природного тела.
Почва — это слой наземных биогеоценозов, где происходит разложение, минерализация и гумификация органического вещества. В ней встречаются все стадии разложения животных и растительных остатков: опавшие листья и начинающие гнить листья и корни растений, микроорганизмов. Все это резко расширяет спектр пищевых ресурсов видов и создает возможность одновременного сосуществования в почве животных с различными пищевыми предпочтениями.
Не менее значительным фактором, определяющим видовое богатство почвенной фауны и огромной ее биомассы по сравнению с обитателями других ярусов биогеоценозов, считается то, что животные используют дополнительный резерв белка — из микроорганизмов почвы, а не только от высшей растительности. И здесь не столь важны общие запасы органического вещества, сколько большие ресурсы доступного белка (Д. А. Криволуцкий, А. Д. Покаржевский).
По степени связи с почвой различают три основные группы животных:
- геобионты - проводящие в почве всю жизнь: дождевые черви, некоторые виды многоножек, ногохвосток и др;
- геофилы - у которых какая-то часть цикла развития обязательно проходит в почве: жужелицы, хрущи, комары-долгоножки и др;
- геоксены - случайные обитатели почвы, использующие почву лишь в качестве временного убежища или укрытия: развивающиеся вне почвы пауки, вредная черепашка и др.
Таким образом, широта условий жизни в почве делает ее средой, переходной между водной и наземной. Животные заселили подстилку и минеральные горизонты почвы благодаря специальным адаптациям к различным ее фазам. Расхождение в образе жизни различных размерных групп привело к формированию различных экологических групп — от физиологически водных до строго наземных. Разнообразие источников пищи также обусловило высокий уровень численности, разнообразие видов и экологических групп. Значение почвы в эволюции животного мира заключается в том, что почва рассматривается как среда, через которую животные могли перейти от водного образа жизни к наземному (М. С. Гиляров).
Глава 2. Трофическая структура почвенных беспозвоночных
Комплекс почвенных беспозвоночных включает разные функционально-ценотические группы, различающиеся как по типу питания, так и по форме деятельности. По типу питания выделяются группы:
- фитофаги — животные, питающиеся живыми растительными тканями;
- зоофаги — животные, питающиеся другими животными. К ним относятся хищники и паразиты;
- сапрофаги — животные, питающиеся разлагающимися остатками организмов;
- миксофаги – формы со смешенным питанием.
Наиболее характерной частью почвенного комплекса являются сапрофаги. На их долю приходится основная масса почвенных животных. Биогеоценотическая роль сапрофильного комплекса состоит как в непосредственном биохимическом и физическом возодействии на органические остатки, так и в стимуляции деятельности сапрофитного комплекса (М. С. Гиляров, Ю. И. Чернов). На основе собственного материала и данных мировой литературы Б. Р. Стригановой проведен подробный анализ питания сапрофагов и раскрыты основные особенности детритных пищевых цепей в почве. Комплекс почвообитающих сапрофагов неоднороден по характеру питания входящих в его состав животных. В нем выделяются трофические группировки: фитосапрофаги, микробофаги (микрофитофаги), детритофаги (Б. Р. Стриганова).
Фитосапрофаги утилизируют непосредственно отмершие ткани сосудистых растений. Эти животные способны использовать клетчатку, сапробиотических нематод, гемицеллюлозы, пектины. Наиболее активными разрушителями листового опада являются диплоподы, мокрицы, наземные моллюски, некоторые виды дождевых червей и коллембол, личинки типулид и бибионид. В переваривании грубой растительной пищи у этих форм активную роль играют кишечные симбионты — бактерии, грибы и простейшие — хотя они имеют собственный фермент целлюлазу.
Микробофаги (микрофитофаги) — это потребители бактериальных пленок, микромицетов, почвенных водорослей. К ним относятся многие виды панцирных и тироглифодных клещей, энхитреид, нематод, коллембол, простейших. Животные отличаются наличием специальных энзимов, расщепляющих оболочки и включения грибных клеток. В этой группе имеются потребители почвенных водорослей — альгофаги, как разновидность фитофагов. Но в основном потребление водорослей комбинируется с бактерио- и микофагией. Совместная встречаемость беспозвоночных и водорослевых группировок имеет большое значение в формировании пионерных сообществ в развивающихся почвах (И. В. Стебаев). При выветривании горных пород на рыхлых продуктах их разрушения в первую очередь поселяются водоросли, лишайники и беспозвоночные — альгофаги (микроартроподы). Экскременты животных составляют основу органической фракции первичных почв. Водоросли благодаря водоудерживающей способности создают среду, благоприятную для поселения микроорганизмов (Э. А. Штина, М. М. Голлербах).
Детритофаги потребляют растительные и животные остатки, утратившие исходную структуру и перемешанные с почвой. Эта группа включает дождевых червей, энхитреид, орибатид, личинок некоторых видов жуков и двукрылых, диплопод семейства полизониида, коллембол.
Среди беспозвоночных почвы, потребляющих энергию разлагающихся органических остатков, М. С. Гиляров выделил такие экологические группы, как сапрофаги (детритофаги), микофаги, копрофаги. Копрофаги — питаются собственными экскрементами или других животных (жуки-навозники, личинки серых мясных мух). К сапрофагам в широком смысле слова относят некрофагов — потребителей трупов животных (жуки-мертвоеды, личинки синих и зеленых мясных мух, кожееды). В почве много видов со смешанным типом питания - миксофагов.
Детрит — это измельченные, сильно разложившиеся и гумифицированные остатки животных и растений с населяющими их живыми микроорганизмами, часто перемешанные с частицами почвы. Он представляет последнюю стадию трансформации органического материала. Пищевая цепь, начинающаяся с детрита, называется «детритная пищевая цепь».
Детритная пищевая цепь почвенного яруса экосистемы играет огромную роль в потоке энергии. Согласно многочисленным литературным данным доля потребляемой первичной продукции фитофагами изменяется в зависимости от биоклиматических условий в пределах 2—30% в различных экосистемах. Таким образом, основная масса первичной продукции (98—70%) при отмирании растений поступает в детритный цикл разложения и наряду с продуктами метаболизма и трупами фитофагов и хищников служит источником энергии и элементов минерального питания животным-сапрофагам и микроорганизмам. Большое количество органического материла поставляют в почву и отмирающие корни растений. Схема детритной пищевой цепи на примере экосистемы широколиственного леса приведена на рисунке 1.
Левый вертикальный ряд – это органические остатки в почве на разных стадиях разложения: опад; непереваренные остатки, прошедшие ферментативную и микробиальную обработку в кишечнике наземных и почвенных животных; детрит. В среднем ряду – трофические группировки организмов сапроблока: сапрофитная микрофлора, первичные (фитосапрофаги) и вторичные разрушители (копрофаги относятся к этому блоку).
Особенность почвенной пищевой цепи заключается в том, что продукты биогенной переработки органических остатков одними формами служат пищей для других групп. Хотя многие сапрофаги являются полифагами, но обладают пищевой специализацией в отношении степени предварительного разрушения растительных остатков (механической и химической деструкции органического материала). Фитофаги же, в отличие от сапрофагов, специализированы к биохимическим особенностям видов или групп кормовых растений, у которых прижизненные выделения оказывают аттрактивное или репелентное влияние на животных (Б. Р. Стриганова).
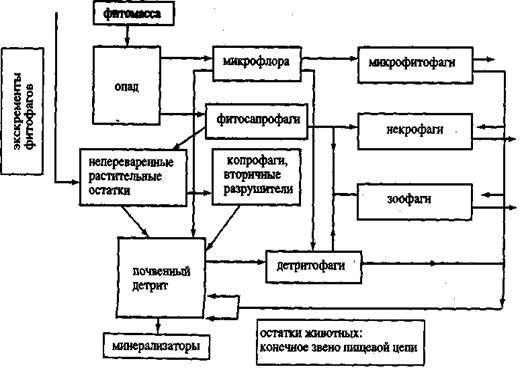
Рисунок 1. Схема детритной пищевой цепи в почве (по Б. Р. Стригановой, 1980)
Среди почвенных сапрофагов по их избирательности к разлагающимся органическим остаткам немецкий ученый В. Дунгер выделил первичных и вторичных разрушителей.
К первичным разрушителям он отнес крупных почвенных и подстилочных беспозвоночных, питающихся отмершей растительностью, но не разложившейся, сохранившей тканевую структуру. Однако растительные остатки начинают перевариваться ими после выщелачивания полифенольных соединений или предварительного разрушения. Животные размельчают и мацерируют частицы растительных тканей в ротовой полости и кишечнике и разрушают связи между отдельными клетками. Они могут быстро размельчать и заглатывать мягкие ткани гниющей древесины, особенно влажной. Совершая вертикальные миграции, беспозвоночные заносят органические остатки в своих кишечниках в глубокие слои и выбрасывают их, формируя копрогенную массу. Этим они стимулируют деятельность сапротрофных микроорганизмов. Существенно то, что первичные разрушители способны переваривать структурные компоненты остатков высших растений — клетчатку, гемицеллюлозы и пектины (Б. Р. Стриганова). Но сапрофитофаги быстрее разрушают «хрупкие» виды листвы нежели остатки с сильно кутикулизированными покровами (корни, кора, листва). К активным разрушителям растительных остатков относятся красные дождевые черви, диплоподы, мокрицы, личинки длинноусых двукрылых (типулиды, бибиониды, ликорииды), некоторые виды коллембол и орибатид.
Доступность и глубина трансформации растительных остатков зависит от механической прочности растительных тканей, содержания трудно разлагающихся веществ — клетчатки, лигнина, пектина и ряда токсичных веществ, угнетающих деятельность микроорганизмов и животных, скорости их выщелачивания (дубильных веществ, танинов и пр.). Так, например, установлено, что буковый опад в широколиственном лесу разлагается очень медленно, слабо потребляется животными. Причиной этого явления оказалось то, что выщелачивание полифенольных соединений из букового опада идет медленно, хотя содержание их не выше, чаи в других породах. В этих условиях первичными разрушителями являются почвенные грибы.
К вторичным разрушителям относятся животные – потребители мертвых растительных остатков, размельченных в кишечнике первичных разрушителей, частично переваренных энзимами животных и микроорганизмов, и обогащенных продуктами их обмена. Они усваивают легкогидролизируемые продукты разложения растительных тканей. Эта категория включает формы со смешанным питанием: сочетание таких пищевых режимов, как сапрофагия, микрофагия, хищничество на мелких животных. Эти беспозвоночные участвуют в процессах деструкции, регулируя состав сапрофитной микрофлоры и способствуя смене грибной фазы разложения микробиальной. Среди почвенных микрофитофагов превалируют потребители микромицетов (Б. Р. Стриганова).
К этой группе относятся копрофаги и детритофаги (дождевые черви, навозники, бронзовики). Значительная доля приходится на мелкие формы (микроартроподы, энхитреиды), обитающие в ферментативном слое подстилки и ризосфере растений, обогащенных разложившимися органическими остатками. Сапрофагия как комплекс пищевых режимов животных показан в таблице 1.
Основной вывод: в комплексе почвенных беспозвоночных - сапрофагов наблюдается последовательный переход к разным пищевым режимам. Наиболее древним пищевым режимом, вероятно, была неизбирательная детритофагия. Черты наибольшей биологической примитивности среди почвенных сапрофагов имеют детритофаги, потребители бактерий, водорослей, простейших. Возможно, на ранних этапах развития почвенного покрова на Земле растительные остатки разрушались микрофлорой.
Таблица 1
Пищевые режимы у беспозвоночных сапрофильного комплекса (по Б. Р. Стригановой)
| Микрофитофаги (микробофаги) | Фитосапрофаги | |
| Первичные разрушители | Вторичные разрушители | |
| Бактериофагия Микофагия Альгофагия | Сапроксилофагия | Копрофагия |
| Сапрофиллофагия | Детритофагия | |
| Сапроризофагия | Потребление жидких продуктов разложения | |
Специализация питания почвенных животных шла по пути формирования микофагии и хищничества и, наконец — сапрофагии. Сапрофаги, переваривающие структурные элементы растительных клеток (первичные разрушители), и фитофаги — эволюционно наиболее молодые группы (Б. Р. Стриганова).
Зональная изменчивость комплекса почвообитающих животных имеет ряд тенденций. Общая численность и биомасса их увеличиваются от тундр к широколиственным лесам и луговым степям и снижаются в направлении к аридным областям. В каждой природной зоне складывается свой специфический комплекс животных. Наиболее бедные сапрофильные комплексы характерны для холодных пустынь и тундры, где преобладают микрофитофаги и практически отсутствуют фитосапрофаги. Первичные разрушители появляются в лесотундре и достигают максимума в подзоне широколиственных лесов. В степях обилие и таксономическое разнообразие комплекса сапрофагов уменьшается, но имеются все функциональные группировки. В аридных условиях значение детритных цепей снижается и увеличивается поток энергии через цепи выедания: здесь происходит сочетание сапро- и фитофагии у животных.
Таким образом, особенности субстрата и состав его потребителей определяют специфичность процесса деструкции не только в разных экосистемах, но и в пределах одной экосистемы. Это говорит о том, что для установления реальной картины участия почвенных беспозвоночных в трансформации органических остатков необходимы исследования конкретных экосистем в конкретных условиях.
Глава 3. Деятельность беспозвоночных животных в почвообразовании
Основные функции животных в разложении растительных остатков следующие (Д. А. Криволуцкий):
а) Пропуская через кишечник большую массу растительных тканей, животные размельчают их и тем самым многократно увеличивают суммарную поверхность растительного материала, доступную микроорганизмам, а также для воздействия воздуха и воды.
б) С помощью собственных ферментов и энзимов симбиотических микроорганизмов беспозвоночные расщепляют целлюлозные компоненты клеток и высвобождают лигнин, который находится в сложном соединении с клетчаткой, что имеет большое значение для развития процессов гумификации органических остатков в почве.
в) В ходе пищеварения в кишечнике почвенных беспозвоночных имеет место частичная минерализация растительных остатков, а у некоторых групп — и частичная гумификация. Экскременты животных — одна из составляющих почвенного гумуса.
г) Многие почвенные животные заглатывают вместе с органическими пищевыми веществами минеральные частицы почвы, способствующие перетиранию в кишечнике пищи. Минеральные частицы (глинистые, песчаные) перемешиваются в кишечниках, спрессовываются и склеиваются выделениями кишечника, образуя разной величины «структурные отдельности» почвы — зернистые комочки; чем выше их количество, тем плодороднее почва.
д) Совершая вертикальные миграции в почве, животные заносят растительные остатки в глубокие горизонты и перемешивают органические и минеральные частицы. Передвижения животных способствуют улучшению условий аэрации почвы, что, в свою очередь, стимулирует аэробные процессы разложения органических остатков.
Исследования круговорота элементов в наземных сообществах показывают, что большое значение в питании беспозвоночных имеет азот и его соединения (А. Д. Покаржевский, Д. А. Криволуцкий). Но внесение азотных минеральных удобрений не оказывает положительного эффекта на численность животных. Это связано с тем, что животные используют в пищу не минеральные формы азота, а органические его соединения — белки, витамины. Питательная ценность белков зависит от их аминокислотного состава. Животные не способны синтезировать незаменимые аминокислоты: лизин, гистидин, аргинин, треонин, валин, метионин, изолейцин, лейцин, фенилаланин, триптофан. Их достаточно только в животном белке и в белках микроорганизмов. Незначительное количество аминокислот в почве и растительных остатках свидетельствует о том, что сапрофаги могут испытывать белковый голод. Существуют два пути преодоления дефицита аминокислот: хищничество и использование микробиальной массы экосистемы. Дефицит белка восполняется и копрофагией. Известны случаи «автокопрофагии» у мокриц и диплопод (Б. Р. Стриганова): животные потребляют растительный опад, который переваривают в очень небольшой степени, а также собственные экскременты, полежавшие в почве и заселенные микрофлорой, которая активно разрушает лигно-целлюлозный комплекс. В экскрементах животные усваивают легкоусвояемые продукты микробного распада структурных компонентов растительных тканей – это основной энергоресурс. Недостаток белковой пищи удовлетворяется за счет микробной массы. Автокопрофагия характерна для животных, у которых отсутствует комплекс облигатных симбионтов в кишечнике.
Почвенные беспозвоночные имеют симбиотические отношения с разными группами микрофлоры — аммонификаторами, фиксаторами молекулярного азота и разрушителями клетчатки. Перерабатывая растительные остатки в пищеварительном тракте животного, они обеспечивают его необходимыми аминокислотами. Но в пищеварительной системе может развиваться и не симбиотическая микрофлора, поступающая с пищей. Очевидно, поглощение почвы тоже связано с потреблением микроорганизмов.
Азот и фосфор для различных групп беспозвоночных — лимитирующие элементы. Азот в виде белковых тел является конструктивным материалом живых организмов биосферы, фосфор в виде молекул органических фосфатов является преобразователем энергии в биосфере. Связь между этими элементами реализуется благодаря деятельности микроорганизмов в экосистемах суши.
С деятельностью различных групп животных почвоведы связывают образование различных форм гумуса (W. Kubiena A. Brauns и др.).
Грубый гумус-мор. Отличается кислой реакцией, малым количеством экскрементов животных. Содержит неразложившиеся и слабо разложившиеся грубые растительные остатки, пронизанные гифами грибов. Отсутствуют прочные связи органических веществ с минеральной глинистой составляющей. Такой гумус образуется во влажном и прохладном климате, когда опад богат трудно разлагаемым материалом — лигнином и целлюлозой, как под хвойными породами. В формировании такого перегноя участвуют раковинные амебы, клещи-орибатиды, ногохвостки. Участие животных незначительно, преобладает грибной тип разложения. Мор свойственен преимущественно подзолистым и дерново-подзолистым почвам.
Мягкий гумус-мулль. В его структуре заметны лишь мелкие неразложившиеся растительные фрагменты. Это результат интенсивной деятельности энхитреид, дождевых червей, многоножек, мокриц, личинок насекомых. В кишечнике животных органическое вещество растительных остатков претерпевает сложные превращения и перемешивается с минеральными частицами. В результате образуется органо-минеральный комплекс. Особенно заметна роль крупных дождевых червей, которые, вовлекая растительные остатки с поверхности в глубь почвы и перемешивая их с минеральными частицами почвы, способствуют созданию зернистой структуры и углублению аккумулятивного горизонта. Кроме того, накапливаются водопрочные экскременты разных животных. Эта форма гумуса присуща серым и коричневым лесным почвам под лиственными лесами и черноземам.
Промежуточная форма гумуса-модер (волокнистый гумус) имеет умеренно кислую реакцию. Он образуется благодаря жизнедеятельности мелких членистоногих, экскременты которых смешиваются с минеральными частицами. В нем умеренное содержание разложившихся фрагментов растительности: структура измельченных частей еще различима, и они не пронизаны гифами грибов. Лучше всего эта зоогенная форма гумуса образуется в условиях повышенной влажности, которая складывается в нижних влажных слоях подстилки под более свежим опадом. В этих условиях в переработке подстилки принимают участие и крупные беспозвоночные.
Но нужно иметь в виду, что в одинаковых растительных сообществах могут формироваться различные типы гумуса. Это зависит от различий в характере почвенной фауны, микрофлоры, абиотических факторов. Так, в буковых лесах на северо-западном Кавказе, где очень бедное почвенное население, отсутствуют черви, подстилка медленно разлагается, слеживается в виде плотного толстого пласта, прочно переплетенного грибным мицелием. Образуется грубый перегной. А в буковых лесах Закарпатья почва богата членистоногими и червями, подстилка быстро разлагается, образуется мулль (Г. Ф. Курчева).
Взаимосвязь компонентов органического комплекса почвы: корневые системы — беспозвоночные — гумус установлена М. С. Гиляровым (1965). Содержание и распределение гумуса по почвенному профилю коррелирует с вертикальной миграцией различных видов беспозвоночных, их численностью, биомассой, пищевой избирательностью и активностью.
Рассмотрим почвообразовательную деятельность отдельных групп беспозвоночных, населяющих разные типы почв, в т. ч. и юга России.
Дождевые черви. Большинство дождевых червей относится к семейству люмбрицид (Lumbricidae). Черви встречаются в разных природных зонах. Они отсутствуют в полярных пустынях, а в полупустынях и пустынях, в неокультуренных зональных почвах. Могут обитать лишь в оазисах или приречных незаселенных почвах. Наибольшей плотностью и наибольшим видовым разнообразием люмбрицид характеризуются зоны смешанных и широколиственных лесов, лесостепь и интразональные луговые ценозы. К югу от лесостепи зоомасса дождевых червей начинает снижаться. В черноземных почвах степей распространены только собственнопочвенные формы дождевых червей и то только в почвах плакоров в очень небольшом числе и только в западных районах степной зоны. Но большая численность может быть в интразональных биотопах (в почве лугов, речных террас, на склонах оврагов) и вблизи жилья, в антропогенных биотопах (на огородах, в компостах и т. п.).
По характеру используемой пищи среди дождевых червей выделяют два морфо-экологических типа. Первый тип — виды, питающиеся на поверхности почвы слабо разложившимися растительными остатками и даже зелеными частями травянистых растений (первичные разрушители по Дунгеру). Например, Lumbricus terresticus. Второй тип — собственнопочвенные черви, питающиеся перегноем почвы. Например, Nicodrilus caliginosus. Представители второго типа значительно энергичнее перемешивают и рыхлят почву, чем первого.
По характеру вертикального распределения по профилю почвы поверхностнообитающие формы (первый тип) делят на три группы: подстилочные — в деятельном состоянии населяющие подстилку и не мигрирующие даже при пересыхании или промерзании почвы глубже 5—10 см; почвенно-подстилочные черви, мигрирующие глубже 10—20 см лишь при неблагоприятных сезонных условиях; норниковые — имеющие постоянно глубоко уходящие (на 1 м и более) ходы, которые они обычно не покидают. Собственнопочвенные дождевые черви (второй тип) не выходят на поверхность почвы при питании или спаривании. Они могут быть разделены на две группы: роющих — обитателей глубоких горизонтов, откуда они мигрируют вглубь только при неблагоприятных гидротермических условиях; норников — имеющих постоянные глубокие ходы, но питающихся также в гумусовом горизонте почвы.
Дождевые черви способствуют минерализации и гумификации растительных остатков, стимулируют деятельность микрофлоры. У них большая потребность в азоте. Предполагается, что ограниченность запасов азота в минеральных горизонтах почвы стала причиной перехода червей к питанию мало разрушенными растительными остатками. Благодаря прохождению пищи через кишечник червей и усилению развития микрофлоры в их экскрементах почва обогащается фульво- и гуминовыми кислотами. Они повышают подвижность азота, калия, фосфора, магния, кальция. При высокой численности содействуют нейтрализации кислых почв: их известковые железы выделяют кальций в кишечный тракт, где воздействуют на пищу. Дождевые черви обогащают почву витаминами группы В, РР, белками: они содержатся в организме животных и после гибели их выделяются в окружающую среду.
Благодаря размножению микрофлоры в кишечнике червей происходит ускорение разложения органики по сравнению с почвой. В экскрементах червей ускоряется минерализация органики: там резко возрастает численность актиномицетов и других микроорганизмов. Дождевые черви способствуют развитию простейших (жгутиконосцев, инфузорий, саркодовых) в почве и увеличению их разнообразия. В целом дождевые черви положительно влияют на биологическую активность почв.
Несомненный интерес представляют диплоподы (Diplopoda), или двупарноногие многоножки. Они играют большую роль в процессах первичного разложения и минерализации органических остатков. В подавляющем большинстве это фитосапрофаги, т. е. питаются опавшей листвой, ветками, гниющей древесиной, предварительно размельченными в ротовой полости. Высокая усвояемость пищи у диплопод объясняется высокой активностью целлюлазы в кишечнике. Собственная целлюлазная активность усиливается ферментами симбиотических микроорганизмов. В кишечнике происходит мацерация растительных тканей. Кроме того, многие виды регулярно потребляют собственные экскременты. При дефиците влаги охотно переходят к фитофагии. Питание живыми растениями восполняет дефицит влаги в организме. Но в силу этого в агроценозах диплоподы становятся вредителями сельскохозяйственных культур. Высокая пищевая активность, способность к потреблению слабо разрушенного опада, высокая усвояемость пищи делают их важнейшим агентом почвообразования (С. И. Головач).
Очень важна роль диплопод в круговороте кальция (Ю. Б. Бызова). Они накапливают кальций в скелете и теле. Получая кальций с пищей, диплоподы в течение жизни выделяют его в виде простых соединений с экскрементами, личиночными шкурками и при отмирании, обогащая им поверхностный слой почвы. Диплоподы могут быть индикаторами почв с высоким содержанием кальция.
Диплоподы широко распространены в различных природных зонах. Наиболее предпочитаемым местом обитания является лесная подстилка широколиственных лесов. Их встречаемость и обилие уменьшаются в почвах открытых ландшафтов — лугов, степей, полупустынь, пустынь и обработанных земель. Они отсутствуют в зоне тундр.
Диплоподы обитают в опаде и верхних горизонтах почвы, уходя в периоды покоя на глубину до 30 см или в гнилую древесину. Они чувствительны как к недостатку, так и избытку влаги в среде: не употребляют излишне увлажненную, с капельками воды, пищу. Нормально двигаются и питаются при хорошей аэрации подстилки, поэтому избегают уплотненных нижних слоев мощной подстилки, где идут гнилостные процессы.
Весьма обычны диплоподы в степной зоне европейской части страны. Кивсяк— характерный элемент сапрофильного комплекса почвенных беспозвоночных в различных биогеоценозах степной зоны Нижнего Дона. Мозаичность почвенно-растительного покрова и, соответственно, гидротермического режима обусловливают пестроту пространственной структуры вида. Кивсяки заселяют байрачные и пойменные леса, овражно-балочные лесонасаждения, лесополосы, заросли кустарников и степные участки. На юго-востоке Ростовской области в полупустынных ландшафтах этот вид отсутствует. Контрастность показателей численности и биомассы свидетельствует, что он предпочитает лесонасаждения и естественные леса. Большое значение играет природный состав насаждений: кивсяки проявляют пищевую избирательность к различным видам лесного опада и ветоши. Это определяет темпы разложения растительных остатков и характер почвообразовательных процессов. Этот вид кивсяка может быть использован для оптимизации процесса разложения в условиях изменяющейся экологической обстановки в разных ландшафтах юга России.
Жуки-чернотелки — характерный и массовый элемент аридных биогеоценозов. Это организмы — минерализаторы, которые производят глубокую переработку опада, препятствуя его трансформации в гумус, сдвигая его в сторону минерализации. Кроме непосредственного поедания подстилки, чернотелки разлагают подстилку, измельчая ее и увеличивая тем самым поверхность разлагающегося субстрата. Экскременты чернотелок — центры стимуляции целлюлозоразрушающей микрофлоры, что дополнительно усиливает разложение подстилки. В ряду зональных степных биогеоценозов уровни численности и биомассы чернотелок увеличиваются от северного предела степей к сухим, после чего снова уменьшаются.
Глава 4. Материалы и методы проведения исследований
Похожие работы


... последствиях изменения среды и сделать лишь косвенные выводы об особенностях самого фактора. Таким образом, при оценке состояния среды желательно сочетать физико-химические методы биологическими. Актуальность биоиндикации обусловлена также простотой, скоростью и дешевизной определения качества среды. Например, при засолении почвы в городе листья липы по краям желтеют еще до наступления осени. ...

... масштабах вряд ли целесообразно, так как поверхностно-активные вещества сами загрязняют среду и появится проблема их сбора и утилизации (Пиковский, 1993). 3.2 Основные подходы и роль биоремедиации в восстановлении нефтезагрязненных почв Существующие механические, термические и физико-химические методы очистки почв от нефтяных загрязнений дорогостоящи и эффективны только при определенном ...
... индикации находят также применение при селекции устойчивых линий лесных, плодовых и декоративных деревьев (Николаевский, 1979; Dassler, 1981 а). 3. Действие антропогенных стрессоров на характер распространения и динамику популяций беспозвоночных животных Количество исследований по воздействию газообразных промышленных выбросов на животных резко возросло именно в последнее десятилетие, после ...
... (металлизация биосферы). Появление новых трансурановых химических элементов, развитие ядерной техники и энергетики. Выход за пределы планеты, развитие космонавтики. 8. С/х ЭС в условиях техногенеза. Агроэкосистема (АЭС) – совокупность биогенных и абиогенных компонентов участков суши преобразованных человеком, используемых для производства сельхозпродукции. Основа АЭС – почва, с/х угодия. ...
0 комментариев