Навигация
3. Семейство генов Wcs 120
Озимые, по сравнению с яровыми злаками, обладают эффективными механизмами акклиматизации, которые позволяют им перезимовывать и выживать при температуре замерзания почвы. Это различие генетически запрограммировано и включает в себя сложную генетическую систему. Чтобы понять характер данной системы и ее регуляцию низкими температурами, были идентифицированы и охарактеризованы гены пшеницы, устойчивой к замерзанию почвы. В ходе этих исследований установлено, что семейство гена wcs120 кодирует группу белков с молекулярными массами от 12 до 200 кД. Как показано при помощи биохимических, иммуногистохимических и молекулярно-генетических анализов, данное семейство генов, специфическое у Poaceae, весьма многочисленно и координированной регулируется низкими температурами. Более того, накопление белков WCS прямо коррелирует с уровнем устойчивости растений к замерзанию почвы. Эти анализы обнаружили также регуляторны контроль процесса яровизации при экспрессии генов низкой температуры у озимых злаков.
Индуцируемые низко температурой у пшеницы гены семейства wcs120 были картированый с использованием вестерн- и Саузерн-блоттинга на дителоцентрических сериях сорта Chines Spring. Идентифицированные гены были локализованы в длинных участках гомологичных групп 6-х хромосом всех трех геномов гексаплоидной пшеницы. Близкие виды, также несущие A, АВ или D геномы также исследовались с использованием вестерн- и Саузерн-блоттинга с wcs120- зондами и WCS120- антителами. Все близкородственные виды несли один или более геномов гексаплоидной пшеницы и продуцировали 50 кДа белок, реагирующи с антителами и определяемы при помощи вестерн-блоттинга, и wcs120-гомологи, которые определялись при помощи Саузерн-блоттинга во всех видах. Отсутствие участка 6DL хромосомы в гексаплоидной пшенице сорта Chines Spring вызывало отсутствие синтеза 50 кДа белка, что не только указывало на локализацию wcs120 в данном участке 6DL-хромосомы, но также и подтвердило молчание wcs120-гомологов в 6А хромосоме. Были также исследованы содержание белков, реагирующих с антителами на WCS120, и уровни холодоустойчивости в сериях лини с заменой хромосом сорта Chines Spring на хромосомы сорта Cheyenne. В ходе экспериментов было установлено, что 5А хромосома сорта Cheyenne увеличивала холодоустойчивость пшеницы сорта Chines Spring. Денситометрия вестерн-блоттингов для определения содержания белка показала, что хромосома 5А имеет регуляторный эффект на экспрессию гена семейства wcs120, располагающегося на хромосомах 6-о группы всех трех геномов гексаплоидной пшеницы.
4 Специфические для низкой температуры и генных богатых глицином белков
Специфическая для низко температуры кДНК пшеницы pTACR7, представляет ген, определенный как tacr7 из озимой пшеницы. Термин низкотемпературно-специфический используется авторами, поскольку экспрессия гена tacr7 не вызывается обработкой экзогенно абсцизовой кислотой или другими типами стресса, такими, как солевой стресс, обезвоживание и тепловой стресс. PTACR7 был выделен при помощи олигонуклеотидного зонда, сходного с pHVCR8 из ячменя посредством RT-PCR с мРНК из ткани побегов пшеницы. На основании предсказанной аминокислотной последовательности были определены характеристики этого белка. Установлено, что TACR7 очень гидрофобный белок, с единственно трансмембранно областью и богат лейцином. Изучение уровне содержания копи tacr7 показало их накопление в проростках пшеницы, верхушечной ткани и каллусной культуре после переноса объектов из контрольных условий на холод. В ходе экспериментов наблюдалось отсутствие обнаруживаемых нозерн-гибридизацие транскриптов pTACR7 в проростках или каллусах, обработанных экзогенно абсцизовой кислото, после солевого стресса, обезвоживания или теплового стресса. Транскрипты tacr7 накапливались в ткани меристемы во время экспонирования при 20C в больше степени в холодоустойчиво озимо пшенице 16029), чем холодочувствительной 16169) с наибольшим различием между генотипами на третьей неделе закаливания. Таким образом, кодируемы tacr7 белок по своим характеристикам уникален среди описанных в литературе индуцируемых низко температурой белков пшеницы.
При изучении мутации сердцевины пентамера CCGAC двух предполагаемых индуцируемых низко температурой элементов в 5'-области индуцируемого холодом гена BN115 озимого рапса при помощи анализа нерезидентной экспрессии равнодействующих мутированных слияний BN115-промотор-GUS было установлено уменьшение низкотемпературного регулирования промотора. Это указывает на то, что последовательность CCGAC в гене BN115 является чрезвычайно важно для его ответа на низкую температуру. Напротив, мутация двух G-боксов CACGTG, находящихся между индуцируемыми низко температурой элементами в то же области промотора, не изменила индуцируемую холодом экспрессию гена. Замена возможно области энхансера промотора BN115 на энхансер из промотора CaMV 35S выразилась в увеличении уровня низкотемпературно индукции экспрессии GUS.
При изучении кДНК ячменя были получены ген-специфические зонды, кодирующие два типа глицин-богатых белков: HvGRP2, характеризуемы цитокератин-подобно и цистеин-богато областями, и HVGRP3, чья основная особенность - наличие РНК-связывающей области. В ходе исследования было установлено, что экспрессия генов HvGRP2 и HVGRP3, которые присутствуют в одно или двух копиях на кажды гаплоидны геном, была повсеместно, и было показано, что ген HVGRP3 модулируется изменением услови освещения. Холодовое возде ствие также увеличивало уровни содержания мРНК HvGRP2 и HVGRP3.
При изучении действия окружающей среды на экспрессию индуцируемых холодом генов на уровне изменения содержания мРНК и тестировании взаимосвязи экспрессии генов и холодно акклиматизации растений ячменя в фазе третьего и четвертого листа было проведено несколько вариантов опыта. В первом варианте растения были выращены в различных температурных условиях между 20/150C и +4/-40С, во втором варианте растения были перенесены с температуры 20/150C на температуру 6/20C и в третьем варианте опыта растения были выращены в условиях засухи или недостатка питательных веществ. В ходе экспериментов измерялись при помощи метода отрастания морозоустойчивость растений и уровни содержания мРНК для трех индуцируемых холодом генов: blt4.9, blt14 и blt101 из меристематических тканей побегов. Результаты экспериментов показывают, что закаленность растени и уровни экспрессии мРНК генов blt4.9, blt14 и blt101 повышались при более низких температурах роста. В ходе экспериментов было также установлено, что на степень изменения содержания мРНК этих генов в ответ на изменение температуры среды значительное влияние оказывали предшествующие температурные условия и возраст растения. При этом уровень закаленности растений сильно коррелировал с уровнями содержания мРНК этих генов в растениях, выращенных в различных температурных условиях. Эта корреляция не распространялась на растения, подвергнутые действию недостатка питательных веществ или засухи.
При изучении влияния низко температуры на экспрессию генов в Poncirus trifoliata из закаленных к холоду растений были клонированы шесть кДНК, представляющие уникальные индуцируемые холодом последовательности. В ходе экспериментов было обнаружено, что гены pbcorc115 и pbcorc119 принадлежат к одному семейству генов. Данные сиквенса указывают на то, что pbcorc115 и pbcorc119 содержат открытую рамку считывания, кодирующую полипептид с молекулярно массой 19.8 кДа и полипептид с меньшей молекулярной массой 11.4 кДа, соответственно. Анализ предсказанной аминокислотной последовательности обнаружил три больших повтора в COR19 и только один повтор в COR11. В каждом повторе были выявлены два элемента -Q-кластеризованный тракт и K-богатый мотив. K-богатый мотив был сходен с подобными мотивами у белков класса D-II из хлопка и группы 2 белков LEA. Сериновый кластер, характерный для многих сходных с группой 2 LEA белков, также был найден в этих белках. В то же время было обнаружено, что в них он находился в необычном положении относительно карбокси-конца. В COR19 и COR11 также присутствовал двусторонни мотив основных остатков, сходных с известными ядерными маркерными последовательностями, что позволяет предположить, что члены данного семе ства белков могут иметь ядерную маркерную функцию. В ходе экспериментов авторами была исследована экспрессия мРНК COR19 в ответ на холодовую акклиматизацию, засуху, затопление и засоление. Результаты показали, что экспрессия COR19 в ткани листа индуцировалась в ответ на холодовую акклиматизацию, но подавлялась во время засухи и затопления.
W. Goodwin с соавторами при изучении озимого рапса изолировали кДНК и выделили ген, кодирующий гибридный богаты пролином белок. Предполагаемы белок по структуре является модульным. При анализе его аминокислотной последовательности установлено, что N-терминальная область данного белка имеет свойства сигнального пептида, который может направлять белок в эндоплазматический ретикулум. Последовательность аминокислот от 27 до 287 имеет три области, которые содержат высокие уровни содержания пролина и нескольких других аминокислот, часто встречающихся в богатых пролином белках клеточной стенки. Эти области характеризуются повторяющимся аминокислотным мотивом. С-терминальная область содержит три предполагаемых мембранно-связывающих участка и имеет высокую степень сходства с аминокислотами некоторых разновидностей гибридных богатых пролином белков. На основе этих данных авторы делают вывод, что данный белок секретируется из клетки, размещается в клеточной стенке и заякоривается в плазматической мембране посредством С-терминально области. В ходе экспериментов установлено, что транскрипты, кодирующие данный белок, индуцируются в ткани листа в течение 8 ч обработки холодом и их содержание быстро снижается при возвращении растения к нормальным температурам. В то же время установлено, что транскрипты не индуцируются тепловым шоком, обезвоживанием, экзогенно абсцизовой кислотой или раневым стрессом, тогда как транскрипты контрольного гена B. napus индуцируются обезвоживанием и экзогенно абсцизовой кислотой.
При изучении экспрессии генов кукурузы были выделены кДНК и соответствующий геномный клон члена семейства генов, кодирующих a-субъединицу фактора элонгации трансляции 1. Предсказанная аминокислотная последовательность из 447 остатков прерывается в гене одним интроном. Саузерн-блот анализ показал, что клонированный ген ef1a - один из семейства, состоящего, по меньше мере, из шести генов. Как показал нозерн-блот анализ, содержание мРНК данного белка в листьях кукурузы увеличивается при низко температуре, в то время как в корнях уровень мРНК ef1 о резко уменьшается. Эти результаты показывают, что экспрессия EF1 о дифференцированно регулируется в листьях и корнях во время холодового стресса.
Похожие работы
... . Эти данные также позволяют предполагать, что некоторая часть белков, индуцируемых холодом и абсцизовой кислотой, связана с солевым стрессом. 2. Влияние гипотермии на содержание водорастворимых белков в тканях бактерий и водорослей Изменение экспрессии водорастворимых белков в ответ на понижение температуры наблюдается также и у водрослей и у бактерий. Во время резкого понижения температуры ...
... различных варианта влияния: усиление холодовой закаленности и усиление формирования каллуса во время регенерации криоконсервированных концов побегов. Белки, препятствующие льдообразованию Одной из функций белков, синтезирующихся в растениях при гипотермии, в частности, при действии отрицательных температур, является препятствование процессу льдообразования. Хотя, как было отмечено выше, в ...
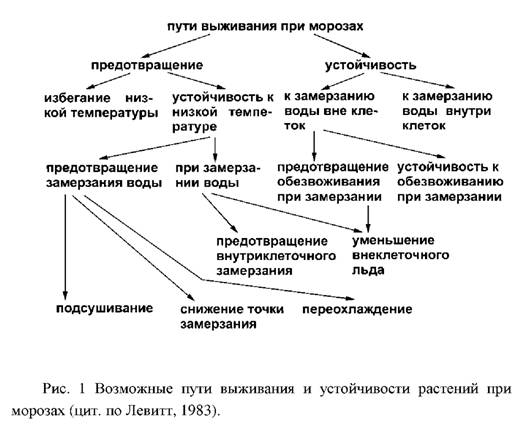
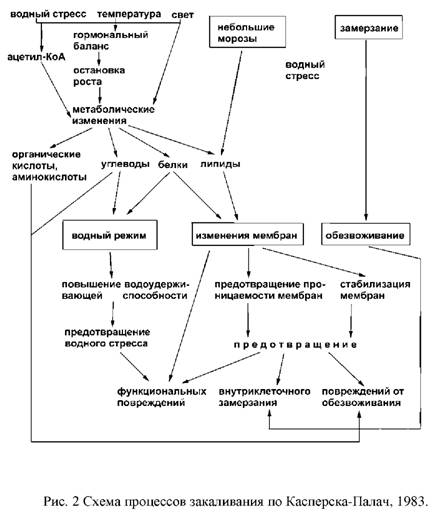
... и выбранные для контроля скорости роста при низко температуре и степени зимнего закаливания, были значительно менее устойчивы, чем норвежские популяции. Степень раззакаливания растени увеличивалась с повышением температуры. В условиях действия низко температуры северный экотип из Bodo был более устойчив к раззакаливанию, чем AberHerald. Тем не менее, при 18°C абсолютны уровень раззакаливания у ...
... Все созданные до сих пор гипотезы, касающиеся вопроса о том, в каком виде хранится запоминаемая информация, можно подразделить на две основные категории. Согласно одной, основой памяти является синтез новых макромолекул, не существовавших ранее в организме, а в соответствии с другой, более обоснованной экспериментально, — субстратом памяти служит формирование устойчивых взаимодействий элементов ...
0 комментариев