Навигация
Гены белков, связанных с процессом льдообразования
55732
знака
0
таблиц
0
изображений
6. Гены белков, связанных с процессом льдообразования
Установлено, что бактериальный ген, связанны с образованием льда, inaZ при перемещении в трансгенные растения вызывает образование ядер льда. Предварительная инкубация преобразованной ткани растения при температурах около 0 0C существенно повысила активность нуклеации льда в растениях, хотя максимальная активность нуклеации льда была достигнута только после низкотемпературно обработки продолжительностью около 48 часов. Хотя трансгенные растения содержат сходные количества мРНК inaZ как при «нормальных», так и при низких температурах, установлено, что низкие температуры необходимы для накопления белка INAZ. Предлагается, что устойчивость INAZ белка и, таким образом, активность нуклеации льда в трансгенных растениях усиливается при низкотемпературной обработке.
7. Гены, кодирующие белки, связанные с передачей кальциевых сигналов
Вопрос о механизме восприятия и трансформации сигнала в клетках растений к настоящему времени все еще изучен недостаточно. Установлено, что роль вторичного посредника в передаче стрессовых сигналов у различных организмов, в том числе и у растений, играют ионы Ca2+. В клетках растений, не подвергнутых действию стресса, уровень содержания Ca2+ при помощи активных Ca2+-транспортирующих систем поддерживается на низком, наномолярном уровне. Холодовой шок вызывает резкое увеличение концентрации Ca2+ в цитоплазме. На животных клетках во время стресса были показаны активации ионами Ca2+ протеинкиназы С и Ca2+-кальмодулинзависимо протеинкиназы, вызывающие изменения экспрессии генов. Для выяснения роли Ca2^ процессах ответа растения на низкотемпературны стресс и адаптации к низко температуре были проведены эксперименты по изучению влияния хелатора Ca2+ - ЭГТА и блокаторов кальциевых каналов - La3+ и верапамила - на процессы адаптации растени люцерны к холоду. Результаты экспериментов показывают, что ЭГТА ингибирует низкотемпературную адаптацию на 70%, а La3+ и верапамил полностью блокируют развитие процесса низкотемпературно адаптации у люцерны.
Чтобы далее исследовать путь передачи сигнала, который стимулирует ответ на холодовой шок у кукурузы, был выделен кодирующий низкотемпературно-индуцируемую кальций -зависимую протеинкиназу клон кДНК. Эксперименты по изучению динамики ответа на стресс показали, что низкотемпературная индукция Zmcdpk1 предшествует аналогичному показателю для mlip15, другого холодоиндуцируемого гена, кодирующего ДНК-связывающий белок типа ле цин-зиппер, что указывает на то, что Zmcdpk1 может размещаться раньше mlip15 в пути передачи вызванного холодным стрессом сигнала. При этом было отмечено, что уровень mlip15 мРНК в конститутивном состоянии в большой степени возрос после обработки циклогексимидом. Кроме гена mlip15, циклогексимид увеличивает уровни транскрипции двух других индуцируемых низко температуро генов -Zmcdpk1 и Adh1, кодирующего алкогольдегидрогеназу 1. Напротив, экспрессия гена хальконсинтетазы индуцировалась только низко температуро. Аккумуляция транскриптов mlip15 при низких температурах и в ответ на циклогексимид значительно уменьшалась после предварительно обработки хелатором кальция, что позволяет предполагать, что кальций вовлечен в обоих случаях индукции гена mlip15.
После холодового шока усиливается экспрессия TCH-генов из Arabidopsis, которые кодируют калмодулин-связанные белки и ксилоглюкан эндотрансглюканазу. Была исследована возможная роль колебаний во внутриклеточных концентрациях иона кальция Ca2+ в вызванно холодовым шоком экспрессии гена TCH. Для этого у трансгенных растени, несущих ген апоэкворина, был проведен мониторинг содержания Ca2+ с тем, чтобы исследовать необходимость индуцируемого холодом увеличения содержания Ca2+ для экспрессии TCH. Индуцированное холодовым шоком увеличение содержания Ca2+ может быть заблокировано La3+ и Gd3+, предполагаемыми ингибиторами Ca2+-канала в плазматической мембране, и 1.2-бис этан-н,н-тетраацетиловой кислотой, внеклеточным хелатором Ca2+. В ходе экспериментов было установлено, что индуцируемая холодовымшоком экспрессия TCH генов затормаживается высокими уровнями содержания всех этих агентов, что, как было показано, блокирует увеличение содержания Ca2+. Эти данные подтверждают то, что внутриклеточное увеличение содержания Ca2+, следующее за холодовым шоком, требует внеклеточного Ca2+ и может получить необходимы приток Ca2+ опосредованно через Ca2+ каналы плазмалеммы. Во-вторых, результаты экспериментов свидетельствуют, что индуцируемая холодом экспрессия хотя бы подкласса TCH-генов требует увеличения содержания внутриклеточного Ca2+.
8. Гены белков холодового шока
Установлено, что у бактерий существует многочисленное семейство генов, экспрессирующихся только в ответ на резкое снижение температуры. Гены этого семейства обозначены как гены белков холодового шока. Показано, что для начала экспрессии некоторых из этих белков холодового шока бактерий достаточно снижения температуры на 10 - 150С.
Известно, что экспрессия гена CspA, важнейшего белка холодового шока Escherichia coli, очень сильно индуцируется во время резкого снижения температуры. Установлено, что замена трех оснований около последовательности Shine-Dalgarno размером в 159-оснований в 5'-нетранслируемой области мРНК cspA стабилизирует мРНК, приводя к конститутивному синтезу CspA при 370С. Как оказалась, эта стабилизация, по крайне мере частично, происходит из-за сопротивления деградации РНКазо E. Холодовая индукция cspA также достигалась путем замены его промотора промотором Ipp, не индуцируемым-холодовым шоком. Таким образом, полученные данные указывают на то, что ген CspA эффективно транскрибируется даже при 370C. В то же время трансляция мРНК белка CSPA заблокирована вследствие ее предельно нестабильности при 370C. Представленные результаты также демонстрируют, что ген cspA конститутивной транскрибируется при всех температурах; однако его экспрессия при 370C предотвращена из-за дестабилизации его мРНК.
С целью оценить эффективность промоторов белков холодового шока для их низкотемпературно экспрессии, было сконструировано транскрипционное слияние генов между промотором гена cspA и геном бета-галактозидазы lacZ. Показано, что в таком слитом гене синтез бета-галактозидазы эффективно подавлялся при 370C, но быстро индуцировался при переносе в 150C, приводя к трех- пятикратному увеличению в специфическо активности белка относительно контрольно бактериально культуры. Только продолжение инкубации культуры при 200C, но не при 150C, приводило к ослаблению активности из-за деления клетки и репрессии промотора, хотя исходные показатели накопления бета-галактозидазы при 200C были в два раза выше измеренных при 150C.
Из двух штаммов Lactobacillus plantarum были клонированы два гена белков холодового шока, cspL и cspP. Эти гены, являющиеся неаллельными, присутствовали во всех исследованных штаммах. Данные гены кодируют полипептиды из 66-аминокислотных остатков, родственные друг другу и принадлежащие к семейству CSP - «белков холодового шока». Транскрипция гена cspP оканчивается единственной мРНК, в то время как для гена cspL были найдены две cspL мРНК с общими 5' концами. Содержание этих транскриптов умеренно возрастало в ответ на обработку культур холодом.
При изучении ответа лактозно-кислых бактерий на замораживание было установлено, что при замораживании культуры лактозно-кислых бактерий жизнеспособность клеток в значительно мере снижалась. В то же время, когда культуры бактерий перед замораживанием были предварительно подвергнуты в течение 2 часов холодовому шоку при 100C, значительно улучшалась жизнеспособность у штаммов Lactococcus lactis subsp.lactis и Pediococcus pentosaceus PO2, но она не менялась у штамма Lactococcus lactis subsp. cremoris и у штаммов Lactobacillus helveticus LB1 и Streptococcus thermophilus TS2. Жизнеспособность клеток возросла еще более, когда период холодового шока был продлен до 5 часов. Использование дегенерированных PCR пра меров, полученных как из гена важнейшего белка холодового шока Escherichia coli, так и Bacillus subtilis приводит к образованию PCR продуктов у всех изученных штаммов. PCR продукт из Lactococcus lactis ssp. Lactis M474 был клонирован и секвенирован. Вычисленная для этого белка последовательность аминокислот показала высокую гомологию с аминокислотными последовательностями других белков семе ства CSP. Использование PCR праймеров, основанных на последовательность M474, приводило к образованию PCR продуктов только у изученных штаммов лактококков, но не у изученных штаммов Lactobacillus helveticus, Streptococcus thermophilus или Pediococcus pentosaceus.
Похожие работы
... . Эти данные также позволяют предполагать, что некоторая часть белков, индуцируемых холодом и абсцизовой кислотой, связана с солевым стрессом. 2. Влияние гипотермии на содержание водорастворимых белков в тканях бактерий и водорослей Изменение экспрессии водорастворимых белков в ответ на понижение температуры наблюдается также и у водрослей и у бактерий. Во время резкого понижения температуры ...
... различных варианта влияния: усиление холодовой закаленности и усиление формирования каллуса во время регенерации криоконсервированных концов побегов. Белки, препятствующие льдообразованию Одной из функций белков, синтезирующихся в растениях при гипотермии, в частности, при действии отрицательных температур, является препятствование процессу льдообразования. Хотя, как было отмечено выше, в ...
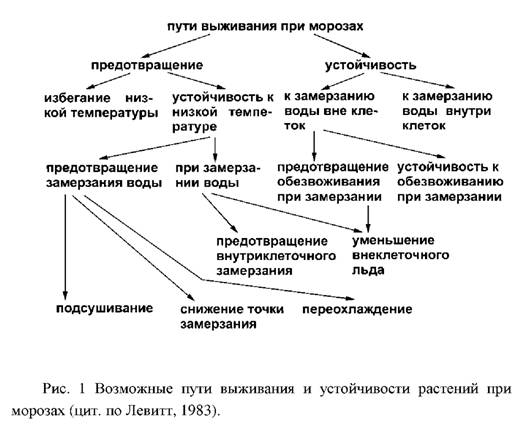
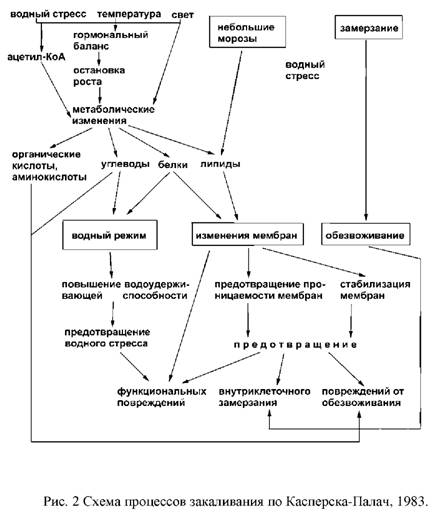
... и выбранные для контроля скорости роста при низко температуре и степени зимнего закаливания, были значительно менее устойчивы, чем норвежские популяции. Степень раззакаливания растени увеличивалась с повышением температуры. В условиях действия низко температуры северный экотип из Bodo был более устойчив к раззакаливанию, чем AberHerald. Тем не менее, при 18°C абсолютны уровень раззакаливания у ...
... Все созданные до сих пор гипотезы, касающиеся вопроса о том, в каком виде хранится запоминаемая информация, можно подразделить на две основные категории. Согласно одной, основой памяти является синтез новых макромолекул, не существовавших ранее в организме, а в соответствии с другой, более обоснованной экспериментально, — субстратом памяти служит формирование устойчивых взаимодействий элементов ...
0 комментариев