Навигация
1.3 ДНК бактерій
Генетичний апарат бактерій представлений молекулою ДНК, що у вигляді нуклеотида розташовується в центральній частині цитоплазми. У вірусів геном представлений ДНК або ж РНК. Молекула ДНК складається з великого числа нуклеотидов. Кожен нуклеотид являє собою з'єднання з фосфату, цукру й азотної підстави. У ДНК входить цукор дизоксирибоза, у РНК - рибоза. У молекулу ДНК вірусів і фагов може входити від декількох тисяч до сотень тисяч нуклеотидов, у ДНК бактерій і найпростіших до 10 млн., а в ДНК вищих організмів до 1 млрд.
Англійський учений Ф. Лемент і американський Дж. Уотсон (1953) обґрунтували представлення про ДНК. На їхню думку молекула ДНК складається їхніх двох ниток, спірально закручених одна навколо іншої. Діаметр подвійної спіралі ДНК дорівнює 2 нм, а відстань між витками 3,4 нм. На кожен виток спирали приходиться 10 пара нуклеотидов. Відстань між азотистими підставами складає 0,34 нм. Її порівнюють із крученими сходами. Якщо згорнуту в спіраль ДНК розгорнути, то вона приймає вид сходів. Цукор і фосфат складають основу подовжніх ниток, "поперечини" складаються з азотистих основ. Азотисті підстави - щільні кільцеподібні з'єднання з атомів С и N (аденин, гуанін, Тимин, цитозин). Ті самі у всіх видів організмів. ДНК різних видів розрізняється порядком чергування зазначених азотистих основ. Як відомо, до складу білків входять 20 амінокислот. Кожній амінокислоті відповідає визначений триплет, тобто три азотистих підстави. Сукупність усіх триплетів одержала назву генетичного коду. Код однаковий у бактерій, вірусів, найпростіших, тварин, людини, тому що послідовність чергування триплетів у молекулі ДНК і молекулі визначеного білка виявляються єдиними в усім органічному світі. У цій єдності будівлі ДНК - найбільша єдність органічного світу. Азотисті підстави в ДНК двох видів:
1. Двокільцеві (пуринові) - аденін, гуанін - 12 ангстрем;
2. Однокільцеві (піримідинові) - тимін, цитозин - 8 ангстрем.
Азотисті підстави нуклеотидів укладені усередині між витками спирали ДНК і з'єднані водневими зв'язками, які потребують строгої парності основ. А саме, Аденін зв'язується з Тіаміном, Гуанін з Цитазином. Чаргафф (1960), а потім радянські вчені А.Н. Бєлозерський і А.С. Спирин показали, що в будь-якій тканині рослин і тварин, у бактеріальній клітині і вірусній частці, вміст молекул аденіну дорівнює вмісту молекул Тиміну, а вміст цитозина - вмісту гуаніну. Це правило нуклеотидних відносин (А + Г/Т + Ц = 1), що містить в основі будівлі всіх ДНК одержало назву по імені автора - правило Чаргоффа. Сума пуринових основ у будь-якій молекулі дорівнює сумі піримідинових основ. Ця закономірність обґрунтована на великій кількості видів організмів. Вона є доказом того, що усередині спіралі ДНК проти кожної пуринової основи знаходиться піримідинове і, навпаки. Згідно правила Чаргоффа аденін одного ланцюга ДНК зв'язаний тільки з Тиміном інший, а гуанін тільки з цитозином. Пара Аденін-Тимін зв'язана двома водневими зв'язками, а гуанін-цитозин - трьома. Така закономірність з'єднання азотистих основ називається комплементарністю, а азотисті основи комплементарними, тобто взаємно доповнюють один одного. Азотисті основи орієнтовані до середини спирали. Для хромосом еукаріот характерна лінійна будова молекули ДНК, у прокаріотів молекула ДНК замкнута в кільце.
Комплементарність азотистих основ у молекулі ДНК складає головну сутність молекулярних основ спадковості і дозволяє зрозуміти, як при розподілі клітки синтезуються тотожні молекули ДНК.
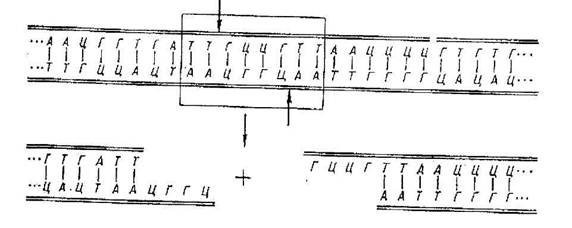
Перед кожним подвоєнням хромосом і розподілом клітки відбувається реплікація (подвоєння) ДНК. Реплікацією називають процес самокопіювання молекули ДНК із дотриманням порядку чергування нуклеотидів, властивим материнським комплементарним ниткам.
Спіралеподібна дволанцюгова ДНК спочатку розплітається (розкручується) уздовж осі, водневі зв'язки між азотистими основами рвуться і ланцюги розходяться. Потім, до кожного ланцюга пристроюються комплементарні азотисті основи й утворюються дві нові дочірні молекули ДНК. Такий спосіб подвоєння молекул, при якому кожна дочірня молекула містить один материнську й один знову синтезований ланцюг, називають напівконсервативним.
Процес реплікації здійснюється за допомогою ферментів, що одержали назва ДНК-полімераз. Ділянка молекули ДНК, у якому почали розплітатися комплементні нитки, називається вилкою реплікації. Вона утвориться в прокаріот у визначеній генетично детермінованій точці. У молекулі ДНК у еукаріот таких точок ініціації реплікації ("стартових точок") буває кілька. У еукаріот процес реплікації ДНК йде неоднаково. Пояснюється це тим, що полінуклеотидні ланцюга в молекулі ДНК антирівнобіжні, тобто 5'-кінець одного ланцюга з'єднується з 3'-кінцем інший, і навпаки. Материнський ланцюг, на якій синтез йде від крапки старту 5'->3' у виді суцільної лінії, називається лідируючої, а другий ланцюг, на якій синтез йде від 3'->5' (у протилежному напрямку) окремими фрагментами одержала назву запізнілої. Синтез цього ланцюга складніше синтезу лідируючого ланцюга. Він протікає за участю ферменту лігази окремими фрагментами. Ці фрагменти (ділянки кодової нитки ДНК) містять у еукаріот 100-200, а в прокаріот 1000-2000 нуклеотидів. Вони одержали назву фрагментів Оказаки.
Фрагмент ДНК від однієї точки початку реплікації до іншої точки утворить одиницю реплікації - реплікон. Реплікація починається з визначеної точки (локус orі) і продовжується доти, поки весь реплікон не буде дуплікований. Молекули ДНК прокаріотичних клітин містять велике число репліконів, тому подвоєння ДНК починається в декількох точках. У різних репліконах подвоєння може йти в різний час або одночасно.
Реплікація молекул ДНК у прокаріот протікає трохи інакше, ніж у еукаріот. У прокаріот одна з ниток ДНК розривається й один кінець її прикріплюється до клітинної мембрани, а на протилежному кінці відбувається синтез дочірніх ниток. Такий синтез дочірніх ниток ДНК одержав назву "обруча, що котиться,". Реплікація ДНК протікає швидко. Так, у бактерії швидкість реплікації складає 30 мкм у хвилину. За хвилину до нитки-матриці приєднується близько 500 нуклеотидів, у вірусів за цей час - близько 900 нуклеотидів. У еукаріот процес реплікації протікає повільно. У них дочірня нитка подовжується на 1,5-2,5 мкм у хвилину.
ДНК усіх живих істот улаштований однаково. ДНК різних видів розрізняються коефіцієнтом видоспецифічності, що являє собою відношення молекулярної суми А + Т к молекулярній торбі Г + Ц. Видоспецифічність ДНК виражається відсотком або часток у ній ГЦ-пара. Коефіцієнт видової специфічності різний у різних видів, але в загальному спостерігається зміна ГЦ-пар від прокаріот до еукаріотам, а в межах останніх - від нижчих до більш високоорганізованих форм.
Вуглеводно-фосфатний кістяк по всій довжині у всіх молекулах ДНК має однотипну структуру і не несе генетичної інформації. Спадкоємна інформація зашифрована різною послідовністю основ. А якщо послідовність основ визначає характер білків собаки, корови, бактерії, вірусу і т.д., те відповідна спадковість може передаватися з покоління в покоління.
Таким чином, у структурній організації молекули ДНК можна виділити первинну структуру - полінуклеотидний ланцюг, вторинну структуру - дві комплементарні один одному полінуклеотидні ланцюги, з'єднані водневими зв'язками, і третинну структуру - тривимірну спіраль з визначеними просторовими характеристиками.
Біологами доведено, що синтез білка відбувається не в ядрі, де локалізована ДНК, а в цитоплазмі. Установлено, що особистої участі в синтезі білка ДНК не приймає. Роль посередника, функцією якого є переклад спадкоємної інформації, що зберігається в ДНК, у робочу форму, виконують рибонуклеїнові кислоти (РНК). Рибонуклеїнова кислота являє собою полінуклеотидний ланцюг, що складається з 4 різновидів нуклеотидів, що містять цукор рибозу, фосфат і одне з 4 азотистих основ - аденін, гуанін, урацил, цитозин. Тому нуклеотиди молекули РНК називаються аденіловою, гуаніловою, урациловою або цитидиловою кислотами. Молекули РНК синтезуються на кодогенного ланцюга ДНК за допомогою Рнк-полімераз з дотриманням принципу комплементарності й антипаралельності. Особливістю є те, що аденіну ДНК у РНК комплементарний урацил. Відомо 3 основних види РНК, що діють у клітині: інформаційна (і-РНК) або матрична, транспортна (т-РНК) і рибосомна (р-РНК).
Інформація про синтез білка з визначеними властивостями укладена в нуклеотидній послідовності матричних або інформаційних РНК (і-РНК, м-РНК), що, у свою чергу, синтезуються на визначених ділянках ДНК. Процес синтезу м-РНК називають транскрипцією. Синтез м-РНК починається з виявлення РНК-полімерази, ділянки в молекулі ДНК, називаного промотором. На цій ділянці Рнк-полімераза розкручує спіраль ДНК і на одній з них фермент синтезує м-РНК. Ланцюг, на якій відбувається зборка молекул м-РНК, називають кодогенною. Зборка рибонуклеотидов у ланцюг відбувається з дотриманням принципів комплементарности й антипаралельності, РНК-полімераза просувається по кодогенному ланцюгу ДНК і здійснює синтез м-РНК доти, поки не зустрічає на своєму шляху термінатор транскрипції (переписування) - специфічну нуклеотидну послідовність. На ділянці розташування термінатора транскрипції Рнк-полімераза відокремлюється від ланцюга ДНК і від синтезованої молекули м-РНК. Промотор (ділянка молекули ДНК), транскретуєма послідовність і термінатора утворять одиницю транскрипції за назвою транскриптон. Після проходження Рнк-полімерази уздовж молекули ДНК, пройдені ділянки поєднуються знову в подвійну спіраль. Утворена матрична РНК містить точну інформацію про білок, записану у визначеній ділянці ДНК. Три поруч розташованих нуклеотидів м-РНК шифрує послідовність амінокислот у пептидному ланцюзі білків. Кожному триплетові (три нуклеотиди - кодони) відповідають визначені амінокислоти. Існує велика розмаїтість і-РНК.
Носієм генетичної інформації бактеріальних кліток є ДНК. Вона являє собою подвійну спіраль, що складається з двох полінуклеотидних ланцюжків. ДНК порівнюють із крученими сходами і з подвійним електричним кабелем. Кістяк ДНК складається з фосфатних груп і дезоксирибози. Поліпептидні ланцюги з'єднані між собою водневими зв'язками, що утримують один з одним комплементарні азотисті підстави. Будівля ДНК бактерій аналогічно такому клітин еукаріотичного типу (рослин, тварин, грибів). На відміну від бактерій у вірусів геном представлений однієї нуклеїновою кислотою - ДНК або РНК. Бактеріальні клітки, крім ДНК, можуть мати генетично повноцінні утворення функціонуючі автономно. Необхідно підкреслити, що носіями спадковості бактерій крім ДНК є плазміди і епісоми. У цьому зв'язку, будь-яка структура бактеріальної клітки, здатна до самореплікації, називається реплікон, тобто репліконами бактерій є нуклеотид, плазміди, епісоми. Плазмиды не зв'язані з нуклеотидом, вони перебувають у цитоплазмі клітки автономно, епісоми можуть знаходитися у вільному стані, але найчастіше вони реплікуються разом із ДНК.
Бактеріальна хромосома представлена однієї двоковою молекулою ДНК кільцеподібної форми і називається нуклеотидом. Довжина нуклеотида в розтягнутому виді складає приблизно 1 мм. Нуклеотид - еквівалент ядра. Розташовано він у центрі бактерії. На відміну від еукаріот ядро бактерій не має ядерної оболонки, ядерця й основних білків (гістонів). Нуклеотид можна виявити у світловому мікроскопі. Для цього треба офарбити клітку спеціальними методами: по Фельгену або по Романовскому-Гимзе. Електронно-мікроскопічне дослідження показало, що один кінець ДНК прикріплений до клітинної мембрани. Видимо, це необхідно для процесу реплікації ДНК.
На відміну від клітин еукаріот у прокаріот відсутні мітохондрії, апарат Гольджи і ендоплазматична сітка.
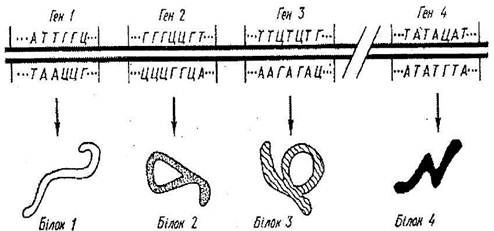
Кожна нитка ДНК складається з ланок - нуклеотидів. До складу нуклеотида входить одна з азотистих основ (аденін, гуанін, тимін або цитозин) дезоксирибоза і фосфорна кислота. Приблизно 1500 нуклеотидів складають ген середньої величини. Таким чином, ген являє собою визначену ділянку ДНК, відповідальний за прояв і розвиток конкретної ознаки. Гени в ДНК розташовані лінійно, вони дискретні, здатні до самореплікації. Послідовність амінокислот у синтезованому білку, визначається послідовністю нуклеотидів у гені.
З погляду функціональної гени підрозділяють на структурні, регулятори, промотори і гени-оператори.
Структурні гени, являють собою гени, що обумовлюють синтез ферментів, що беруть участь у біологічних реакціях і у формуванні клітинних структур.
Гени-регулятори відповідальні за синтез білків, що регулюють обмін речовин. Ці гени можуть впливати на діяльність структурних генів.
Гени-промотори детермінують початок транскрипції. Вони являють собою ділянку ДНК, що розпізнає Днк-залежний Рнк-полімеразою.
Гени-оператори є посередниками між структурними генами, промоторною областю і генами-регуляторами.
Сукупність генів-регуляторів, промоторів, операторів і структурних генів називають опероном. Отже оперон є функціональною генетичною одиницею, що несе відповідальність за прояв визначеної ознаки мікроорганізмів.
Розрізняють індуцибельні і репресибельні оперони. Наприклад, індуцибельним опероном є Lac-оперон, гени якого контролюють синтез ферментів, що утилізують лактозу в мікробній клітці. Якщо клітка не має потреби в лактозі, оперон підтримується в неактивному стані і, навпаки.
Прикладом репресибельного оперона може служити триптофановий оперон, що забезпечує продукцію триптофану. Цей оперон звичайно постійно функціонує, а його білок-репресор знаходиться в пасивному стані. У випадку підвищення змісту триптофану в клітці амінокислота вступає в зв'язок з репресором і активізує його. Репресор інгібує працюючий оперон і перериває синтез триптофану.
Найважливіша властивість ДНК - здатність до реплікації. Реплікація може протікати по тета-типі і сігма-типові. Реплікація ДНК по тета-типі починається у визначеній крапці у виді "здуття" і поширюється уздовж молекули в двох напрямках, проходячи через проміжну структуру, що нагадує грецьку букву тета. При цьому типі реплікації зберігається один з ланцюгів вихідної молекули ДНК, а друга синтезується з нуклеотидів.
Реплікація ДНК по сігма-типі здійснюється через проміжну структуру, що нагадує грецьку букву сигма, відкіля і назва цього типу. Цей тип реплікації спостерігається в процесі кон’югації бактерій і деяких фагів. При цьому типі реплікації відбувається добудовування обох ниток ДНК до дволанцюгової ДНК.
Геном бактерій виконує наступні функції: забезпечує передачу біологічних властивостей; програмує синтез бактеріального білка з визначеними властивостями; бере участь у процесах мінливості бактерій; забезпечує збереження індивідуальності виду; детермінує множинну стійкість до ряду лікарських речовин.
Розділ 2. Генетичні процеси в клітинах мікроорганізмів 2.1 Генетичні рекомбінації у бактерій: трансформація, конюгація, трансдукція
У процесі еволюції природний добір діє не тільки на рівні мутацій в окремих генах, але і на рівні рекомбінації генів, в результаті якої із ДНК двох різних клітин утворюється рекомбінантна хромосома. Цей процес називається генетичною рекомбінацією, а клітини, які утворюються в результаті цього процесу –– рекомбінантами. При рекомбінації не проходитьповне злиття клітин (не утворюються диплоїди), а частина генетичного матеріалу донорної клітини переноситься в реціпієнтну клітину, яка стає частковим диплоїдом або мерозиготою. В рекомбінантній хромосомі основу складає хромосома клітини – реціпієнта, яка включає частину клітини – донора, рекомбінанти формуються в реціпієнтних клітинах.
До рекомбінативної мінливості генетичного матеріалу, приводить трансформація, трансдукція і кон`югація. Рекомбінативна мінливість належить до другого типу спадкової мінливості (після мутаційної).
Тансформація (лат. transformatio – перетворення) передача генетичного від донора до реціпієнта за допомогою ізольованої ДНК. При трансформації не потрібен безпосередній контакт між клітиною–донором і клітиною–реціпієнтом. Джерелом трансформуючої ДНК може служити вбита культура бактерій (свіжа), або чисті препарати ДНК, які з неї екстраговані.
Явище тренсформації у бактерій вперше спостерігав Ф. Гриффітс в 1928 р., але він його не пояснив. Пізніше в 1944 р. О.Евері, К.Мак-Леод і М. Мак-Карті вдалося виділити трансформуючий агент і встановити його хімічну природу. Трансформуючим агентом виявилася ДНК. Процес трансформації проходить у декілька етапів: 1) адсорбція трансформуючої ДНК на поверхні клітини реціпієнта; 2) проникнення ДНК в клітину; 3) з`єднання трансформуючої ДНК з відповідним фрагментом хромосоми реціпієнта.
Не всі клітини бактерій здатні сприймати ДНК. Клітини, які сприймають трансформуючу ДНК, називаються компетентними.
Явище трансформації виявлино у стафілококів, бацил, бульбочкових бактерій, агробактерій, бруцел та ін. Число таких видів перевищило 50, але найкраще вона вивчена у сінної палички і стрептококу пневмонії. Вчені вважають, що можлива і міжвидова трансформація, але це спостерігається тоді, коли ДНК донора і реціпієнта подібні за нуклеотидним складом–гетеротрансформація. Гомотрансформація – перенесення генетичної інформації від одного штаму бактерій до другого (в межах одного виду). Відкриття явища трансформації дало змогу встановити роль нуклеїнових кислот як носіїв спадкової інформації.
За характером розміщення перенесених ознак розрізняють зчеплену трансформацію – перенесення двох і більше генів, які розміщені поруч, одним фрагментом ДНК.
Незчеплена тренсформація – перенесення генів різними фрагментами ДНК, або одним, Але гени не розміщені поруч.
В результаті трансформації утворюються трансформанти, які мають ознаки донора і реціпієнта. Рекомбінантна ДНК далі реплікується як єдина структура. Трансформація може здійснюватися як в лабораторних умовах так і в природі. Трансформацію в бактерій використовують для проведення гібридологічного аналізу різних мутацій, для встановлення філогенетичної подібності донора і реціпієнта.
Кон’югація (лат. conjugatio спряження бактерій) – передача генетичного матеріалу від однієї клітини до другої шляхом безпосереднього контакту між ними.Вперше була вивчена в 1946 р. Дж. Ледербергом і Е. Татумом при культивуванні кишкової палочки.
Пізніше було показано, що між кон’югуючими клітинами утворюється цитоплазматичний мостик і втановлено наявність статевої деференціації. При кон’югації одна бактерія є донором — чоловіча клітина F+ (анг. fertility — плодючість), друга — реціпієнтом — жіноча клітина F—.
Статева диференціація зумовлена статевим фактором (F фактор), який є лише в чоловічих клітинах. Статевий фактор — це дволанцюгова ДНК, яка має форму кільця. Вона зумовлює ряд властивостей чоловічих клітин — наявність статевих ворсинок F-пілі, спецефічну чутливість клітин-донорів до “чоловічих” дрібних РНК і ДНК-вмісних фагів. За допомогою статевих ворсинок чоловіча клітина прикріпляється до жіночої і через їх канальці відбувається перенесення генетичного матеріалу. Якщо схрещувати між собою жіночі клітини, то рекомбінанти не утворюються.
Статевий фактор може існувати в клітині автономно (поза хромосомою). Його відносять до групи бактеріальних плазмід. Але поряд з цим, існують клітини, в яких статевий фактор інтегрований з хромосомою. Такі плазміди дістали назву епісом — і клітини називаються Hfr – клітинами (висока частота рекомбінації — 10 —3).
Під час кон’югації при передачі ДНК від донора до реціпієнта зберігається цілісність генома донорної клітини. В клітину реціпієнта переноситься одноланцюгова ДНК донора під впливом якої в клітині синтезується комплементарний ланцюг і відновлюється дволанцюгова ДНК. Завершується кон’югація утворенням рекомбінантної бактеріальної хромосоми.
Кон’югація може відбуватися між штамами одного виду, між представниками різних видів. Це приводить до утворення так званих міжвидових рекомбінантів.
Трансдукція — процес перенесення генетичного матеріалу від однієї бактеріальної клітини до другої за допомогою бактеріофага. Відкрив в 1952 р. Н. Ціндер і Дж.Ледерберг на прикладі двох штамів сальмонел.
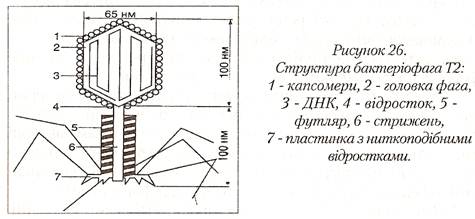
Трансдукцію здійснюють помірні фаги та їх вірулентні мутанти.Суть трансдукції полягає в тому, що деякі помірні фаги в процесі репродукції включають у свій геном невеликі фрагменти ДНК бактерії-донора і переносять їх до бактерій-реціпієнтів.
Фаги діляться на вірулентні і помірні. Вірулентні фаги, проникають в клітину, зумовлюють формування нових фагів і лізис бактерій. Зараження клітин помірними фагами не завжди супроводжується лізисом бактерій, частина їх виживає і стає лізогенними. В лізогенних бактеріях ДНК фага включається в ДНК клітини і помірний фаг перетворюється в профаг, який втрачає здатність руйнувати бактеріальну клітину. Профаг поводить себе так, як частина бактеріальної хромосоми і відтворюється в її складі протягом декількох поколінь.
У бактерій розрізняють 3 типи трансдукції: загальну, специфічну і абартивну.
При загальній трансдукції проходить передача різних фрагментів ДНК від бактерій-донорів до бактерій-реціпієнтів за допомогою помірних трансдукуючих фагів.
Специфічна трансдукція характеризується здатністю фага переносити від бактерій донорів до бактерій реціпієнтів тільки певні гени. Це зумовлено тим, що утворення трансдукуючого фага проходить в результаті з’єднання його ДНК із строго визначеними бактеріальними генами, розміщеними на хромосомі клітини донора.
При абортивній трансдукції перенесений фагом фрагмент хромосоми клітини-донора не включається в хромосому клітини-реціпієнта, а розміщується в її цитоплазмі автономно. В процесі поділу клітини-реціпієнта трансдукований фрагмент ДНК-донора може передаватися тільки одній із двох дочірних клітин, тобто успадковується однолінійно, в зв’язку з чим втрачається в потомстві.
Похожие работы
... і всіх розв'язних документів, необхідна певна законодавча база, що дозволяє у випадках появи негативних наслідків з юридичної точки зору вирішити проблему, що створилася. 1. Теоретичні основи генно-модифікованих продуктів Генетична інженерія - це нова галузь молекулярної біології, яка розробляє методи перенесення генетичного матеріалу від одного живого організму до іншого з метою ...
... живлення, дихання, ріст і розвиток, розмноження, реакції на зовнішні подразники, пластичність, інтенсивність взаємодії з факторами середовища. Фізіологія являється науковою основою промислового використання мікроорганізмів у мікробіологічних виробництвах біологічно активних речовин (БАР), ферментів, вітамінів, антибіотиків, амінокислот, органічних кислот. Мікроорганізми здатні до синтезу ...
... . Основний механізм резистентності Haemophilus influenzae до β-лактамних антибіотиків полягає в продукуванні β-лактамаз, які гідролізують амінопеніциліни та цефалоспорини І покоління. Решта β-лактамних антибіотиків, як правило, зберігають високу активність по відношенню до цього збудника. За даними дослідження PROTEKT, проведеного в 20 країнах світу в 2001-2006 рр., у середньому ...
... » Початок маркування трансгенів в Україні перенесли на 1 грудня, з 1 листопада Кабмін повинен був зобов’язати українських виробників маркувати продукти харчування, які містять генетично модифіковані організми (ГМО) у кількості, що перевищує 0,9 відсотка, що було зроблено. Також 18 лютого 2009 р. була прийнята та затверджена постанова Кабінету Міністрів України, в якій йдеться про те, що всі ...
0 комментариев