Навигация
Регуляція генної активності
47247
знаков
0
таблиц
0
изображений
2.2 Регуляція генної активності
Функціональна нерівнозначність кліток і зв'язана з нею репресія й активація генів давно привертали увагу генетиків.
Перша спроба пояснити регуляторну активність генів були зв'язані з вивченням гістонних білків. Ще чоловік і жінка Стедман на початку 40-х років нашого століття одержали перші чіткі результати про розходження в хімічній природі гістонних білків. Подальші дослідження показали, що регуляція генної активності набагато більш складний процес, ніж простої взаємодія ділянок генів з молекулами пістонних білків.
Жакоб і Моно розділили гени регуляторної системи на два типи - гени-регулятори і гени-оператори. Автори ввели в генетику нове поняття, визначивши блок структурних генів і керуючий ними оператор як єдину функціональну одиницю - оперон.
В останні роки були отримані дані про наявність ще одного керуючого осередку генної активності-промоторі. Виявилося, що по сусідству з операторною ділянкою, до якого приєднується продукт - білкова речовина репресор, синтезований на гені-регуляторі, мається інша ділянка, що відноситься до членів регуляторній системі генної активності. До цієї ділянки приєднується молекула ферменту РНК- полімерази. У цій промоторній ділянці повинне відбутися взаємне дізнавання унікальної послідовності нуклеотидов у ДНК і специфічній конфігурації білка РНК- полімерази. Від ефективності дізнавання буде залежати здійснення процесу зчитування генетичної інформації з даної послідовності генів оперона, що примикає до промотору.
2.3 Позахромосомні фактори спадковостіДо позахромосомних факторів спадковості відносять плазміди і епісоми, що розташовуються в цитоплазмі клітини. Плазміди не здатні вбудовуватися в нуклеотид бактерії, вони мають власну ДНК, що може самостійно реплікуватися. На противагу плазмідам, епісоми вбудовуються в нуклеотид бактерії і функціонують разом з ним.
Плазміди, не залежно від нуклеотиду, забезпечують здатність до коньюгации, стійкість до антибіотиків і інших речовин. Установлено, що наявність плазмид у клітці не обов'язково, але в теж час їх може бути кілька. Плазміди підрозділяють на кон`югативні (трансмісивні) і некон`югативні (на трансмісивні). Перші - додають клітині властивості генетичного донора, детермінують перенос генетичного матеріалу від клітки донора до клітини реципієнтові, другі - не додають клітині властивостей генетичного донора, не можуть передаватися до клітини реципієнта без наявності факторів переносу.
Розрізняють наступні види плазмід: Соl-фактор - коліциногенний фактор, F-фактор - фактор фертильності, R-фактор - фактор стійкості до лікарських речовин, плазміди біодеградації, плазміди, що кодують фактори вірулентності в мікроорганізмів (Ent, Hly, Sal, K і т.д.)
Col-фактори - це плазміди, що контролюють синтез бактеріоцинів, що володіють здатністю пригнічувати розвиток філіпченкових родинних бактерій. Назва бактеріоциногенів привласнюють з урахуванням виду мікроорганізмів їхній продуцирующих. В даний час відомо, що практично майже всі патогенні бактерії продукують бактеріоцини.
Бактеріоцини кишкової палички називають коліцини, стафілокока - стафілоцини, пневмокока - пневмоцини, вібріона - вібріоцини і т.д.. Краще інших бактеріоцинів вивчені коліцини. Культури кишкової палички, продукуючі коліцини, називають коліциногенами, а чуттєві до них - коліциночутливими. Коліцини - речовини білкової природи. Вони мають здатність ингибировать синтез ДНК, РНК, білка, викликати загибель клітки не порушуючи її цілісності. Коліцини мають летальну ознаку, тобто після їхньої продукції бактеріальна клітка може загинути. Коліцини функціонують аналогічно антибіотикам з вузьким спектром дії, мають властивості ендодезоксирибонуклеаз.
Бактеріальні клітини, що виділяють бактерицини, стійкі до дії гомологічних бактерицинів навколишнього середовища.
F-фактор може функціонувати автономно і може бути в інтегрованому, як епісома, стані. Цей фактор являє собою кільцеву ДНК довжиною 30-32 нм, молекула якої детермінує перенос генетичного матеріалу з клітки донора в клітку реципієнта, синтез полових ворсинок, синтез ферментів, здатність до автономної реплікації і т.д.
R-фактор генетична структура, що забезпечує стійкість до лікарських препаратів. Ця структура несе гени лікарської стійкості (год-гени). Стійкість до одному або декільком лікарським препаратам (антибіотикам) здійснюється за рахунок оперонів і може бути передана шляхом кон’югації і трансдукції.
Плазміди біодеградації відповідальні за використання органічних сполук бактеріями як джерела вуглецю й енергії, за утилізацію ряду цукрів, утворення протеолітичних ферментів.
Ent-плазміди кодують утворення ентеротоксинів у ентеробактерій, Hly-плазміда - синтез гемолізинів у ентеропатогенних мікроорганізмів і стрептококів. Sal-плазміда контролює в псевдомонад використання бактеріями саліцилатів завдяки виробленню призначеного для цієї мети ферменту.
2.4 Використання на практиці досягнень генетики мікроорганізмів
Досягнення генетики мають важливе значення в сільському господарстві, промисловому виробництві і в медицині. Внаслідок утворення індукованих мутантів можна отримати в десятки і сотні разів більше цінних продуктів (антибіотиків, ферментів, вітамінів, амінокислот) в порівнянні з дикими формами мікроорганізмів.
Широкі перспективи відкриває перебудова спадкової природи організмів шляхом генної інженерії.
Очевидно, методом генної інженерії можна буде створити такі бактерії, які втратять хвороботворність, допоможуть виробити імунітет проти багатьох інфекційних захворювань людини і тварин. В промисловості появляться високопродуктивні мікроорганізми, які створюватимуть білки, ферменти, вітаміни, антибіотіки, ростові речовини і інші продукти.
Методом генної інженерії будуть створені рослини, які володіють здатністю до зв’язування молекулярного азоту (шляхом трансплантації гена, який відповідає за фіксацію мол. N2 у клітини вищих рослин).
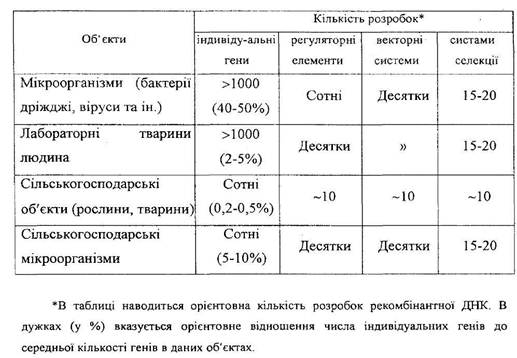
Вивчення генетики бактерій та інших мікроорганізмів має дуже важливе як теоретичне, так і практичне значення для спрямованої селекції високопродуктивних штамів, які останнім часом почали широко застосовуватися у різних галузях народного господарства. Використання в селекції мікроорганізмів методів природного добору, індукованого мутагенезу, популяційної мінливості, клонування, гібридизації соматичних клітин тощо дало можливість одержати високопродуктивні штами мікрорганізмів. Останні знайшли широке застосування в мікробіологічній промисловості для виробництва кормового білка, амінокислот, ферментів, вітамінів, антибіотиків, бактеріальних добрив, засобів захисту рослин, анатоксинів, лікувально-профілактичних препаратів – вакцин, інтерферонів, гормонів, інтерлейкінів та ін. Наприклад, з індукованих мутантів з наступною селекцією їх було одержано штами – продуценти амінокислот, продуктивність яких у сто разів вища від такої у вихідних штамів. Продуцент лізину дає в 300-400 разів більший вихід цієї незамінної амінокислоти, ніж природний штам.
Багатонадійні перспективи для сільського господарста, біології та медицини й інших галузей народного господарста відкриваються у зв’язку з розробкою і вдосконаленням методів генної і клітинної інженерії, за допомогою яких експериментально доведено можливість передачі не тільки природних генів, а й штучно синтезованих, які кодують синтез різноманітних біологічно-активних сполук. Наприклад, ще в перших дослідах з генної інженерії, проведених у 1973 році, було введено за допомогою фага в геном E. coli ген LIG, який контролює синтез лігази. Внаслідок цього вміст лігази в клітинах-реципієнтах збільшився в 500 разів. Тепер у клітини кишкової палички клоновані і функціонують гени інтерферонів, гормону росту, інсуліну та ін. За допомогою клонованих штамів E. coli одержують препарати інтерферону, інсуліну і соматотропіну.
Є також дані про те, що успішно функціонують клоновані у бактерії гени вірусів грипу, гепатиту В, герпесу, ген білка оболонки вірусу ящуру, що в найближчий час дозволить розробити технологію виробництва молекулярних вакцин без баластних білків.
Останнім часом інтенсивно вивчаються методи трансплантації генів за допомогою плазмід, які ще часто називають “генною інженерією у природі”. Вони відіграють велику роль у передачі генетичного матеріалу між бактеріями, які належать навіть до віддалених філогенетичних груп.
Плазміди є фактично каналом генетичної комунікації в бактеріальному світі. Наприклад, методами генної інженерії було зроблено пересадку гена nif з азотфіксуючої бактерії в неазотфіксуючу, і остання набула властивості фіксувати молекулярний азот. Тепер ведуться роботи з перенесення генів від бактерій до клітин вищих рослин.
У лабораторних умовах одержано рекомбінантні плазміди, які містять гени двох різних бактерій, бактерій і вірусів, бактерій і рослин, бактерій і тварин, бактерій і людини. Дуже важливим є те, що такі рекомбінантні плазміди, інродуковані в бактеріальні клітини, дали експресію.
Особливої ваги набувають нині методи одержання енергії та переробки відходів промисловості і сільського господарства з метою одержання цінних біопродуктів і захисту біосфери від забруднення за допомогою мікроорганізмів. Мікробіологічна наука і мікробіологічна індустрія можуть зробити помітний внесок у розв’язання енергетичних проблем, які пов’язані зі значним зменшенням запасів нафти і вугілля на нашій планеті.
Висновки
Найважливішими ознаками живих організмів є мінливість та спадковість. Основу спадкового апарату бактерій, як і всіх інших організмів, складає ДНК. Поряд з цим спадковий апарат бактерій і можливості його вивчення мають ряд особливостей: бактерії - гаплоїдні організми, тобто вони мають 1 хромосому. У зв'язку з цим при спадкуванні ознак відсутнє явище домінантності; бактерії мають високу швидкість розмноження, у зв'язку з чим за короткий проміжок часу (доба) змінюється кілька десятків поколінь бактерій. Це дає можливість вивчати величезні за чисельністю популяції і досить легко виявляти навіть рідкі за частотою мутації. Спадковий апарат бактерій представлений хромосомою. У бактерій вона одна. Якщо і зустрічаються клітини з 2, 4 хромосомами, то вони однакові.
Хромосома бактерій - це молекула ДНК. Довжина цієї молекули досягає 1,0 мм і, щоб "уміститися" у бактеріальній клітині, вона не лінійна, як у еукаріот, а суперспіралізована в петлі і згорнута в кільце.
Генотип (геном) бактерій представлений не тільки хромосомними генами. Функціональними одиницями генома бактерій, крім хромосомних генів, є: ІS-последовності; транспозони; плазміди.
У бактерій розрізняють 2 види мінливості - фенотипову і генотипову.
Фенотипова мінливість - модифікація - не торкається генотипу, але торкає більшість особей популяцій. Модифікації не передаються в спадщину і з часом загасають, тобто повертаються до вихідного фенотипу через більш (тривалі модифікації) або менш (короткочасні модифікації) число поколінь.
Генотипова мінливість стосується генотипу. В ее основі лежать мутації і рекомбінації.
Мутації бактерій принципово не відрізняються від мутацій эукариотических клітин. Особливістю мутацій у бактерій є відносна легкість їх виявлення, тому що існує можливість працювати з великими за чисельністю популяціями бактерій. За походженням мутації можуть бути: спонтанними; індукованими. По довжині: точковими; генними; хромосомними. По спрямованості: прямими; зворотними.
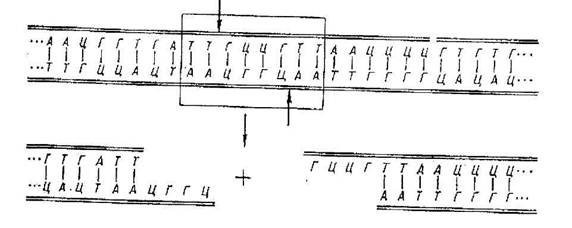
Рекомбінації (обмін генетичним матеріалом) у бактерій відрізняються від рекомбінацій у еукаріот: у бактерій є кілька механізмів рекомбінацій; при рекомбінаціях у бактерій утвориться не зигота, як у еукаріот, а мерозигота (несе цілком генетичну інформацію реципієнта і частина генетичної інформації донора у виді доповнення); у бактеріальної клітини-рекомбіната змінюється не тільки якість, але і кількість генетичної інформації.
Список використаної літератури
1. Атабеков И.Г. Практикум по общей вирусологии. — М.: Из-во Московского университета, 1981. — 191 с.
2. Бойко А Л. Экология вирусов растений. — К.: Вища шк., 1990. — 165 с.
3. Борьба с вирусными болезнями растений. / Под ред. Х.Кеглер. М.: Агропромиздат. — 1986. — 326 с.
4. Вавилов Н.И. Иммунитет растений к инфекционным заболеваниям. - М: Наука, 1986. — 520 с.
5. Векірчик К.М. Практикум з мікробіології: Навч. посібник. – К.: Либідь, 2001. – 144 с.
6. Вершигора А.Ю., Бранцевич Л.Г., Василевская И.А. и др. Общая микробиология. — К.: Вища шк. Головное изд-во, 1988. — 342 с.
7. Віруси і вірусні хвороби бобових культур на Україні. / Московець СМ., Краев В.Г., Порембська Н.Б. та ін. — К.: Наукова думка 1971.— 136 с.
8. Вірусні хвороби сільськогосподарських культур/ Московець С.Н., Бобирь А.Д., Глушак Л.Е. / Під ред. Бобиря А.Д. — К.: Урожай, 1975. — С.72-80.
9. Власов Ю.И. Вирусные и микоплазменые болезни растений. — М.: Колос, 1992. — 207 с.
10. Власов Ю.И., Ларина Э.И. Сельскохозяйственная вирусология. — М.: Колос, 1982. — С. 150-156.
11. Генкель Л. А. Микробиология с основами вирусологии. — М.: Просвещение, 1974. — 270 с.
12. Гиббс А., Харрисон Б. Основы вирусологии растений: Пер. с англ. — М.: Мир, 1978. — 430 с.
13. Гнутова Р.В. Серология и иммунохимия вирусов растений. — М.: Наука, 1993. — 300 с.
14. Гусев М. В., Минова Л. А. Микробиология. М.: Изд-во Моск. ун-та, 1992. - 448 с.
15. Мишустин Е.Н., Емцев В.Т. Микробиология. – М.: Агропромиздат, 1987. – 368 с.
Похожие работы
... і всіх розв'язних документів, необхідна певна законодавча база, що дозволяє у випадках появи негативних наслідків з юридичної точки зору вирішити проблему, що створилася. 1. Теоретичні основи генно-модифікованих продуктів Генетична інженерія - це нова галузь молекулярної біології, яка розробляє методи перенесення генетичного матеріалу від одного живого організму до іншого з метою ...
... живлення, дихання, ріст і розвиток, розмноження, реакції на зовнішні подразники, пластичність, інтенсивність взаємодії з факторами середовища. Фізіологія являється науковою основою промислового використання мікроорганізмів у мікробіологічних виробництвах біологічно активних речовин (БАР), ферментів, вітамінів, антибіотиків, амінокислот, органічних кислот. Мікроорганізми здатні до синтезу ...
... . Основний механізм резистентності Haemophilus influenzae до β-лактамних антибіотиків полягає в продукуванні β-лактамаз, які гідролізують амінопеніциліни та цефалоспорини І покоління. Решта β-лактамних антибіотиків, як правило, зберігають високу активність по відношенню до цього збудника. За даними дослідження PROTEKT, проведеного в 20 країнах світу в 2001-2006 рр., у середньому ...
... » Початок маркування трансгенів в Україні перенесли на 1 грудня, з 1 листопада Кабмін повинен був зобов’язати українських виробників маркувати продукти харчування, які містять генетично модифіковані організми (ГМО) у кількості, що перевищує 0,9 відсотка, що було зроблено. Також 18 лютого 2009 р. була прийнята та затверджена постанова Кабінету Міністрів України, в якій йдеться про те, що всі ...
0 комментариев