Навигация
ФЕДЕРАЛЬНОЕ АГЕНСТВО ПО ОБРАЗОВАНИЮ
ПЕНЗЕНСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ ИМ. В.Г. БЕЛИНСКОГО
Естественно-географический факультет
Кафедра биохимии
КУРСОВАЯ РАБОТА
ПО БИОХИМИИ МЕМБРАН:
«МЕЖКЛЕТОЧНЫЕ КОНТАКТЫ»
Выполнила студентка IV курса группы БХ-41
Гребенникова Ирина
Научный руководитель Соловьев В.Б.
ПЕНЗА, 2009г.
Содержание:
Введение
1.Классификация межклеточных контактов
2. Замыкающие межклеточные контакты
2.1.Рыхлые или простые контакты
2.2.Плотный замыкающий контакт
2.3.Замыкающие межклеточные контакты
3.Адгезионные или сложные межклеточные контакты
3.1.Точечные межклеточные контакты
3.2.Адгезионные пояски
3.3.Адгезионные соединения между клеткой и матриксом
3.4.Десмосомы
4.Гемидесмосома
5.Заболевание аутоиммунное, поражающее десмосомы
6.Проводящие межклеточные контакты
6.1.Синапсы
6.2.Нексусы
7.Рецепторы
Вывод
Приложение 1
Список литературы
Введение
Межклеточные контакты возникают в местах соприкосновения клеток в тканях и служат для межклеточного транспорта веществ и передачи сигналов, а также для механического скрепления клеток друг с другом. Основные типы межклеточных контактов: а) рыхлые, или простые, контакты — между плазматическими мембранами соседних клеток имеется щель шириной 10—20 нм, заполненная гликокалликсом, специализированных структур на мембранах нет; б) межклеточные «замки» — мембраны соседних клеток разделены таким же расстоянием, но изгибаются, образуя на поверхности клеток впячивания; в) десмосомы; г) плотные контакты (встречаются в основном в эпителиальных клетках) — разделяются на зону замыкания и зону слипания (промежуточный контакт); в зоне замыкания две соседние мембраны сливаются своими наружными слоями, эта зона непроницаема для макромолекул и ионов, в зоне слипания мембраны разделены щелью в 10—20 нм, заполненной плотным веществом, вероятно, белковой природы; д) щелевидные (высокопроницаемые) контакты, свойственные всем типам эпителиальной и соединительной тканей,— плазматической мембраны разделены промежутком в 2— 4 нм, пронизанным каналами, по которым низкомолекулярные вещества попадают из цитоплазмы одной клетки в другую, минуя межклеточную среду. В большинстве случаев межклеточные контакты разрушаются при удалении из среды ионов Са2+ . Особыми формами межклеточных контактов являются синапсы, а также плазмодесмы растит, клеток.
1. Классификация межклеточных контактов
1.Замыкающие межклеточные контакты.
а) простой или рыхлый контакт;
б) плотный замыкающий контакт.
2.Адгезионные межклеточные контакты.
а) точечные контакты;
б) адгезионные пояски;
в) адгезионные соединения между клеткой и внутриклеточным матриксом;
г) десмосомы.
3.Проводящие.
а) нексусы;
б) синапсы.
2. Замыкающие межклеточные контакты
2.1 Рыхлые или простые контакты
Простой контакт — соединение клеток за счет пальцевидных впячиваний и выпячиваний цитомембран соседних клеток. Специфических структур, формирующих контакт, нет.
Простые контакты занимают наиболее обширные участки соприкасающихся клеток. Расстояние между билипидными мембранами соседних клеток составляет 15-20 нм, а связь между клетками осуществляется за счет взаимодействия макромолекул соприкасающихся гликокаликсов.
Посредством простых контактов осуществляется слабая механическая связь - адгезия, не препятствующая транспорту веществ в межклеточных пространствах. Разновидностью простого контакта является контакт "типа замка", когда плазмолеммы соседних клеток вместе с участком цитоплазмы как бы впячивается друг в друга (интердигитация), чем достигается большая поверхность соприкосновения и более прочная механическая связь.
2.2 Плотный замыкающий контакт
Плотный замыкающий контакт — соприкасаются билипидные слои мембран соседних клеток. В области зоны плотных контактов между клетками не проходят практически никакие вещества.
Постоянные клеточные контакты скрепляют клетки в эпителиальном клеточном слое таким образом, что предотвращается перетекание даже малых молекул с одной стороны слоя на другую. Латеральная подвижность многих мембранных белков ограничена. Ограничение подвижности достигается с помощью барьеров, образованных при участии плотных контактов.
Клоны эпителиальных тканей (эпителии) функционируют в качестве избирательно-проницаемых барьеров, разделяющих жидкости с разным химическим составом по обе стороны слоя. В выполнении этой функции плотные контакты играют две роли.
Осуществляемый эпителиальными клетками трансклеточный транспорт (например, питательных веществ полости тонкого кишечника во внутриклеточную жидкость по другую сторону слоя) зависит от двух групп мембранных белков-переносчиков: одна находится на апикальной (обращенной в полость) поверхности клетки и активно транспортирует отдельные молекулы в клетку; другая находится на базолатеральной поверхности клетки и позволяет тем же молекулам покидать клетку путем облегченной диффузии . Для поддержания этого направленного транспорта не должно происходить перемещения апикальных белков-переносчиков на базолатеральную поверхность и наоборот.
Кроме того, промежутки между эпителиальными клетками должны быть скреплены таким образом, чтобы транспортированные молекулы не могли бы продиффундировать назад в полость через межклеточные промежутки.
Плотные контакты и выполняют эти две функции: барьеров для диффузии мембранных белков между апикальной и базолатеральной поверхностями и скрепления соседних клеток вместе так, что водорастворимые молекулы не могут перетечь на другую сторону слоя. При этом плотные контакты непроницаемы для макромолекул, а их проницаемость для малых молекул сильно варьирует в разных эпителиях. Эпителиальнные клетки могут временно модифицировать плотные контакты с тем, чтобы допустить увеличенный ток жидкости через бреши в контактных барьерах. Такой параклеточный транспорт особенно важен при абсорбции аминокислот и моносахаридов из полости тонкого кишечника.
Важнейшим элементом в структуре избирательно проницаемых барьеров эпителиальных и эндотелиальных являются плотные контакты. Избирательная проницаемость варьирует от ткани к ткани, пропуская или целые клетки и макромолекулы, или только протоны и ионы. Плотный контакт выглядит как пояс из переплетающихся скрепляющих нитей, который полностью окружает апикальный конец каждой клетки эпителиального слоя. Полагают, что скрепляющие нити состоят из длинных рядов специфических трансмембранных белков в каждой из двух взаимодействующих плазматических мембран, и которые (белки) соединяются напрямую друг с другом, что приводит к закупориванию межклеточного пространства. Интегральным мембранным белком плотного соединения оказался окклудин (взаимодействует с двумя цитоплазматическими белками, ZO-1 и ZO-2 ( zonula occludence 1, 2 ). Их функция окончательно не ясна. Возможно, их роль заключается в локализации оккулдина в сайтах между апикальной и базолатеральной поверхностями клетки. Некоторые ассоциированные с цитоскелетом белки были также обнаружены в участках плотных контактов. Среди них зингулин, антиген и актин (по данным электронной микроскопии, актиновые филаменты состоят из двух цепей глобулярных молекул, диаметром 4 нм и образующих двойную спираль, на каждый виток которой приходится 13,5 молекулы). Эти цепи составляют основу тонких филаментов скелетных мышц, которые кроме актина содержат также несколько других белков; глобулярный актин имеет молекулярную массу около 42 кД. Он содержит одну полипептидную цепь, состоящую из 375 или 374 аминокислотных остатков; различия в аминокислотной последовательности у разных актинов, как в пределах одного вида, так и межвидовые, крайне незначительны. Они составляют не более 25 аминокислотных замен; в настоящее время у позвоночных животных различают 6 изоформ актина, в зависимости от изоэлектрической точки они делятся на 3 класса - альфа, бета и гамма; бета- и гамма-актины характерны для немышечных клеток , а альфа-актины - для мышечных). Ras играет определенную роль в регулировании функционирования плотных соединений. Таким образом, в клетках имеются, по-видимому, сходные механизмы построения и регуляции адгезионных структур, и эти механизмы тесно взаимосвязаны с изменениями в цитоскелете. Однако, каким образом перестройки цитоскелета влияют на процессы межклеточной адгезии, пока окончательно не ясно. Механизмы адгезии и межклеточной сигнализации тесно сопряжены с давно известным феноменом контактного торможения , природа которого до сих пор до конца не выяснена.
Похожие работы
... рассмотренный в тексте обзора межклеточный матрикс. Настоящая работа имеет и ещё один аспект. Рассмотренные в ней данные последних десятилетий показывают, что адекватно понять колониальную организацию и межклеточную коммуникацию микроорганизмов можно лишь в том случае, если учесть всю гамму не только внутривидовых, но и межвидовых экологических отношений. Иначе, говоря биосоциальные микробные ...
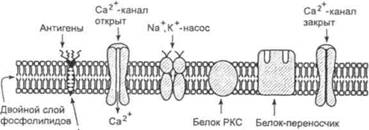
... мембраны; 6) создание иммунной специфичности клетки за счет наличия в структуре мембраны антигенов; 7) обеспечение избирательной проницаемости веществ через мембрану и транспорта их между цитозолем и окружающей средой. Для понимания механизмов гуморальных регуляций в организме необходимо знание не только структуры и проницаемости клеточных мембран для различных веществ, но и структуры и п
... из воды, 9% из органических (6% из них белки - альбумины, глобулины, фибриноген и протромбин) и 1% из неорганических веществ. Рh плазмы около 7,36. Функции крови: 1. Трофические (доставка к тканям питательных веществ). 2. Защитная (фагоцитоз, иммунная защита). 3. Газообмен, т.е. дыхательная функция. 4. Гомеостатическая функция. 5. Интегративная функция (участвует в гуморальной регуляции, ...
... нервами. БИЛЕТ № 12 Общая морфология заднего мозга Задний мозг, включающий продолговатый мозг и Варолиев мост представляют собой филогенетически древнюю область центральной нервной системы, сохраняя черты сегментарного строения. В заднем мозге локализованы ядра и проводящие восходящие и нисходящие пути. По проводящим путям в задний мозг поступают афферентные волокна от вестибулярных и ...
0 комментариев