Навигация
Відведення ЕЕГ проводяться двома способами: монополярным і біполярним
16073
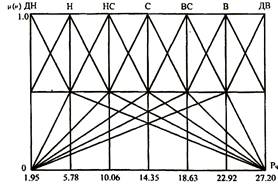
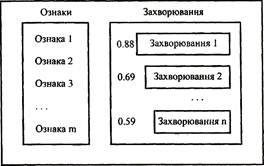
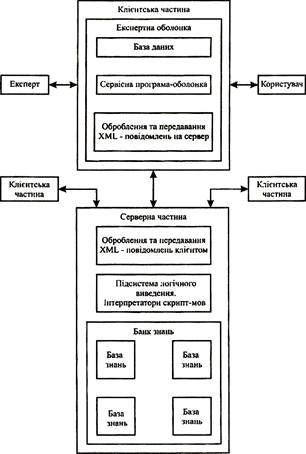
знака
0
таблиц
4
изображения
1. Відведення ЕЕГ проводяться двома способами: монополярным і біполярним
При монополярному способі відведення виміряються потенціали між активною крапкою на голові щодо неактивної зони, що не дає власних ритмів коливань — референтного електрода.
Біполярний метод складається у вимірі різниці потенціалів двох активних зон мозку. Електроди закріплюються на голові за допомогою спеціального шолома чи приклеюють спеціальним клеєм.
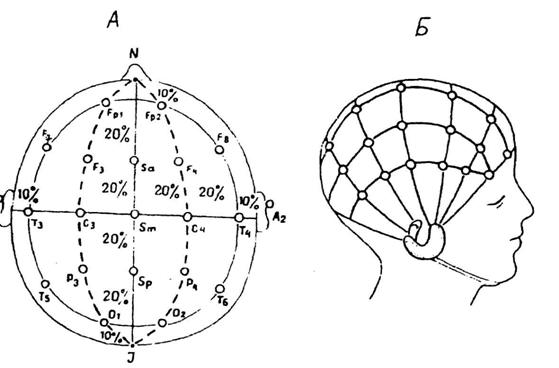
Рис.2. Схема відведень біопотенціалів мозку.(10—20%).
2. ЗОВНІШНЄ ЕЛЕКТРИЧНЕ ПОЛЕ ПІРАМІДНИХ НЕЙРОНІВ КОРИ ГОЛОВНОГО МОЗКУ
Більшість наявних експериментальних даних говорить про те, що генез ЕЕГ (електроенцефалограми) визначається, в основному, електричною активністю кори великих півкуль головного мозку, а на рівні кліток - активністю її пірамідних нейронів. У пірамідних нейронів виділяють два типи електричної активності. Імпульсний розряд (потенціал дії) із тривалістю близько 1 мс і більш повільне (градуальне) коливання мембранного потенціалу — гальмові і збудливі постсинаптичні потенціали (ПСП).
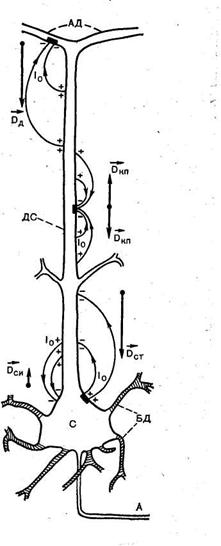
Рис 3. Виникнення токових диполів у пірамідному нейроні нової кори.
С – сома (тіло), А - аксон, БД – базальні дендрити, ДС- дендритний стовбур , ПЕКЛО - дендрити апікального розгалуження (відповідно).
Чорні прямокутники показують деякі ділянки, у яких генеруються ПСП; знаки + і - позначають полярність деяких ділянок зовнішньої поверхні клітки, між якими в зовнішнім середовищі протікає струм І0; Dси – вектор соматичного диполя при імпульсному розряді; Dст, - вектор соматичного диполя при генерації гальмового ПСП; Dд - вектор дендритного диполя при генерації збудливого ПСП; Dкп - вектор диполів квадруполя, що утвориться при генерації збудливого ПСП на дендритному стовбурі.
Гальмові ПСП пірамідних кліток генеруються в основному в тілі нейрона, а збудливі ПСП — переважно в - дендритах. Правда, на тілі нейрона мається визначена кількість збудливих синапсів, і відповідно до цього тіло пірамідних нейронів (сома) здатно генерувати також і збудливі ПСП. Тривалість ПСП пірамідних кліток, принаймні, на порядок більше тривалості імпульсного розряду.
Зміни мембранного потенціалу обумовлюють виникнення в пірамідних клітках двох токових диполів, що відрізняються по цитологічній локалізації (рис. 3). Один з них — соматичний диполь з дипольним моментом Dc. Він формується при зміні мембранного потенціалу тіла нейрона; струм у диполі і в зовнішнім середовищі протікає між сомою і дендритним стовбуром. Вектор дипольного моменту Dc при імпульсному розряді чи генерації в тілі нейрона збудливого ПСП спрямований від соми уздовж дендритного стовбура, а гальмовий ПСП створює соматичний диполь із протилежним напрямком дипольного моменту. Інший диполь, називаний дендритним, виникає в результаті генерації збудливих ПСП на розгалуженні апікальних дендритів у першому, плексиморфному шарі кори; струм у цьому диполі тече між дендритним стовбуром і зазначеним розгалуженням. Вектор дипольного моменту Dд дендритного диполя має напрямок убік соми уздовж дендритного стовбура.
Генерація збудливого ПСП у районі дендритного стовбура без розгалуження приводить до появи квадруполя, оскільки при цьому від частково деполяризованої ділянки струм усередині клітки поширюється в двох протилежних напрямках, у результаті чого формуються два диполі з протилежним напрямком дипольних моментів Dкп (див. рис. 3). Тому що диполі малі в порівнянні з відстанями до крапок відведення ЕЕГ, зовнішнім полем квадрупольного генератора пірамідних кліток можна зневажити.
Важливе значення для з'ясування механізму генеза ЕЕГ має питання про те, який тип електричної активності (потенціал чи дії ПСП) дає найбільший внесок у потенціал зовнішнього поля головного мозку. Ясно, що для генеза ЕЕГ найбільш істотна та електрична активність, при якій виникає диполь з великим дипольним моментом. Величину цього дипольного моменту можна оцінити по довжині позаклітинного поля, наприклад по відстані, на якій потенціал падає до дуже малих значень чи у визначене число раз. К.Ю. Богданов і В.Б. Головчинський розрахували просторовий розподіл позаклітинного електричного поля пірамідних нейронів кори при імпульсній активності і при градуальній зміні мембранного потенціалу соми, обумовленій збудливим ПСП. Цей розрахунок проводився на основі кабельної теорії за умови, що величина потенціалу дії на порядок перевищує ПСП.
По відношенню до потенціалу дії, через її короткочасність, мембрана дендрита поводиться як ємність, що володіє низьким опором струму високої частоти. Тому струм, обумовлений імпульсною активністю, циркулює на невеликій відстані від тіла клітки; ємність мембрани шунтує віддалені ділянки стовбура. Дійсно, за даними мікроелектродних досліджень, зовнішнє електричне поле пірамідних нейронів, яке генерується потенціалом дії, не виявляється вже на відстанях вище 0,1 мм. Таким чином, ЕЕГ повинна в основному створюватися «повільним» соматичним і дендритним диполями, що виникають при генерації гальмових і збудливих постсинаптичних потенціалів.
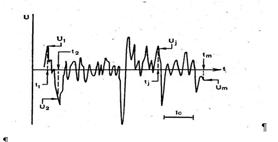
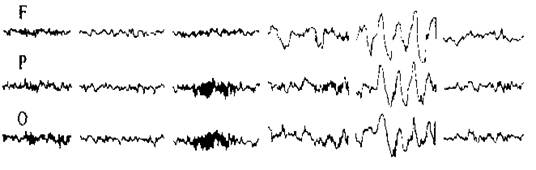
Рис 4. Фрагмент фонової ЭЭГ. U — різниця потенціалів; t — час.
3. СТАНДАРТНЕ ВІДХИЛЕННЯ. ЕЛЕКТРОЕНЦЕФАЛОГРАМИ ЯК КІЛЬКІСНА ХАРАКТЕРИСТИКА ЕЛЕКТРИЧНОЇ АКТИВНОСТІ МОЗКУ
При вивченні зовнішнього електричного поля мозку реєструють і інтерпретують перемінний сигнал ЕЕГ, а постійну складову, як правило, не беруть до уваги. Як видно на рис. 2, ЕЕГ фонової активності мозку представлявляє собою дуже складну залежність різниці потенціалів від часу і виглядає як сукупність випадкових коливань різниці потенціалів. Для характеристики таких хаотичних коливань («шумів») використовують параметри, відомі з теорії імовірності: середню величину і стандартне відхилення ![]() від середньої величини. Щоб знайти
від середньої величини. Щоб знайти ![]() , виділяють ділянку на ЕЕГ, яку розбивають на невеликі рівні інтервали часу, і наприкінці кожного інтервалу (t1, t2, tm на рис. 4) визначають напругу U(U1, Uj,Um на рис.4). Стандартне відхилення
, виділяють ділянку на ЕЕГ, яку розбивають на невеликі рівні інтервали часу, і наприкінці кожного інтервалу (t1, t2, tm на рис. 4) визначають напругу U(U1, Uj,Um на рис.4). Стандартне відхилення ![]() розраховують по звичайній формулі.
розраховують по звичайній формулі.
При вивченні ритмічних ЕЕГ, що характеризуються визначеною амплітудою і частотою зміни різниці потенціалу, показником величини ЕЕГ може служити амплітуда цих коливань.
4. ДИПОЛЬНИЙ ЕКВІВАЛЕНТНИЙ ЕЛЕКТРИЧНИЙ ГЕНЕРАТОР ГОЛОВНОГО МОЗКУ
В даний час у дослідженнях ЕЕГ для моделювання електричної активності кори головного мозку розглядають поводження сукупності токових електричних диполів окремих нейронів. Запропоновано кілька таких моделей, що дозволяють пояснити окремі особливості ЕЕГ. Розглянемо модель М.Н. Жаднюга, що на прикладі генеза ЕЕГ при відведенні з твердої мозкової оболонки дозволяє виявити загальні закономірності виникнення сумарного зовнішнього електричного поля кори.
Основні положення моделі: 1) зовнішнє поле головного мозку в деякій крапці реєстрації - інтегроване поле, яке генерується токовими диполями нейронів кори; 2) генез ЕЕГ обумовлений градуальною електричною активністю пірамідних нейронів; 3) активність різних пірамідних нейронів деякою мірою взаємозалежна (скорелирована); 4) нейрони розподілені по корі рівномірно і їхні дипольні моменти перпендикулярні до поверхні кори; 5) кора плоска, має кінцеву товщину h, а її інші розміри нескінченні; з боку черепа мозок обмежений плоским безкрайнім токонепроводящим середовищем. Обґрунтування перших двох положень розглянуто вище. Зупинимося на інших положеннях моделі.
Дуже велике значення для генеза ЕЕГ має взаємозв'язок електричної активності пірамідних нейронів. Якби градуальна зміна мембранного потенціалу в часі відбувалася в кожнім нейроні зовсім незалежно від інших кліток, перемінна складова потенціалу їх сумарного зовнішнього електричного поля була б невеликою, тому що збільшення потенціалу за рахунок посилення активності одного нейрона значною мірою компенсувалося б хаотичним зниженням активності інших нейронів. Порівняно висока величина региструємої у досвіді ЕЕГ змушує припустити, що між активностями пірамідних нейронів існує позитивна кореляція. Кількісно це явище характеризують коефіцієнтом кореляції RK. Цей коефіцієнт дорівнює нулю при відсутності зв'язку між активностями індивідуальних нейронів і дорівнював би одиниці, якби зміни мембранного потенціалу (дипольних моментів) кліток відбувалися зовсім синхронно. Проміжне значення, що спостерігається в дійсності, Rk свідчить про те, що діяльність нейронів синхронізована лише частково.
Інтегроване поле безлічі диполів-нейронів було б дуже слабким при високому рівні синхронізації, якби вектори дипольних моментів елементарних джерел струму були орієнтовані в корі хаотично. У цьому випадку спостерігалася б значна взаємна компенсація полів індивідуальних нейронів. У дійсності ж, згідно з цитологічними даними, дендритні стовбури пірамідних кліток у новій корі (ці клітки складають 75% від усіх кліток кори) орієнтовані практично однаково, перпендикулярно поверхні кори. Поля, створювані диполями таких однаково орієнтованих кліток, не компенсуються, а складаються.
Таким чином, зовнішнє електричне поле головного мозку, що виявляється у виді ЕЕГ, можна розглядати як результат додавання полів пірамідних нейронів нової кори. Важлива умова такого генеза ЕЕГ — однакова орієнтація диполів нейронів і наявність позитивної кореляції в їх градуальній електричній активності; при цьому достатній навіть дуже низький рівень попарної кореляції.
Похожие работы
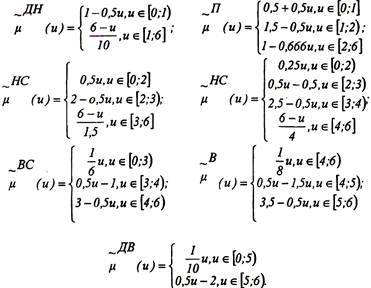
... хребта. Розглянуті технології, мережеві протоколи і програмне забезпечення можуть бути використані для вирішення проблем телемедицини і в інших клінічних областях. Експертна система "Лазерна рефлексотерапія" Застосування експертних систем, призначених для вирішення завдань діагностики і лікування захворювань, особливо ефективне в тих випадках, коли врахування дуже великого обсягу вхідної і ...
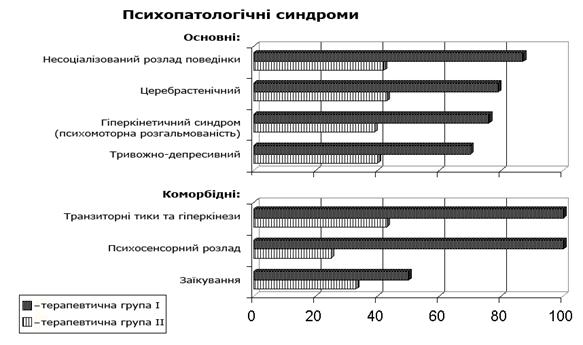
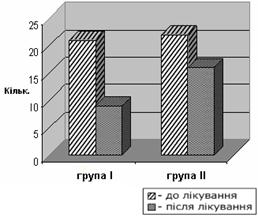
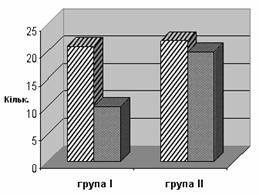
... интоксикацией (ХТИ) // Возрастная нейропсихология и нейропсихиатрия: Материалы научно-практической конференции с международным участием. Киев, 2007 г. – С. 37. Анотація Макаренко С.М. Особливості психічних порушень у дітей з хронічною тонзилогенною інтоксикацією. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.01.16 – психіатрія. – Украї ...
... відповідні збудливі і гальмівні зв’язки здатні генерувати і підтримувати ритмічну активність кори великого мозку. Клінічне значення. Запис ЕЕГ– діагностична процедура, яка використовується в неврологічній практиці. Так, при дифузних органічних пошкодженнях головного мозку, черепно-мозкових травмах спостерігаються сповільнені і нерегулярні хвилі. При пухлинах мозку часто виникають місцеві зміни
... 4 с. (Особисто дисертантом проведено написання тексту, підготовку до друку, подано лист до друку). АНОТАЦІЯ Башкірова Л.М. Клініко-параклінічна характеристика та стан забезпечення макро- та мікроелементами у хворих з пароксизмальними станами при хронічних порушеннях мозкового кровообігу. – Рукопис. Дисертація на здобуття наукового ступеня кандидата медичних наук за спеціальністю 14.01.15 – ...
0 комментариев