Понятие и виды взаимодействия микроорганизмов с высшими растениями
Симбиотические взаимодействия между микроорганизмами и высшим растениям
Влияние фитопатогенных микроорганизмов на высшие растения
Изучение взаимодействия микроорганизмов с высшими растениями на уроках биологии
Практическое применение знаний о взаимодействии микроорганизмов с высшими растениями во внеурочной работе
Навигация
Симбиотические взаимодействия между микроорганизмами и высшим растениям
Взаимодействие микроорганизмов с высшими растениями
73336
знаков
7
таблиц
6
изображений
1.2 Симбиотические взаимодействия между микроорганизмами и высшим растениям
Симбиоз – весьма распространенное явление. Понятие о симбиозе было сформулировано во второй половине XIX века немецким ученым де Бари, который понимал симбиоз в широком смысле слова как взаимосвязи между двумя или несколькими организмами (11, 143).
Симбионты высших растений, представленные микроорганизмами, разнообразны (см. Рис. 3).

Рис. 3 Симбионты высших растений.
Почва представляет собой благоприятную среду для развития ризосферных микроорганизмов. Приблизительные подсчеты показывают, что в 1г почвы содержится 1х109 бактерий, 1х105 грибов, 1х105 актиномицетов, 1х103 водорослей. Суммарная масса всех микробных клеток в пахотном слое составляет примерно 6-7 т на 1 га. Естественно, такое колоссальное количество живых клеток оказывает многообразное и разностороннее влияние на процессы, происходящие в почве, и на жизнь высших растений. (45, 181)
Микрофлора ризосферы, принимая участие в процессах трансформации органических веществ в почве, обеспечивает растения необходимыми элементами минерального питания и некоторыми биологически активными веществами. Кроме того, микроорганизмы ризосферы разлагают многие токсичные для растений соединения, обеззараживая почву.
Микрофлору зоны корня принято подразделять на микрофлору ризопланы – микроорганизмы, непосредственно поселяющиеся на по-верхности корня, и микрофлору ризосферы – микроорганизмы, населяющие область почвы, прилегающей к корню (16, 320). Численность микроорганизмов в ризоплане и ризосфере (Р) в сотни и даже тысячи раз превышает их содержание в окружающий почве (П), что можно выразить отношением: Р/П (см. Приложение 1).
На численность и групповой состав микрофлоры ризопланы и ризосферы оказывает влияние:
·тип почвы,
·климатические условия,
·характер растительного покрова
·стадия развития растений.
Корни растений стимулируют или угнетают микроорганизмов в разной степени. Бобовые растения чаще всего стимулируют развитие микробов. В ризосфере клеверов, например, обнаружено значительно больше микроорганизмов, чем в зоне корней злаков и деревьев.
Корневые выделения растений в случае длительного выращивания одних и тех же культур на одних и тех же площадях приводят к «почвенному утомлению». Такая обстановка в сочетании с одинаковым по составу растительным опадом вызывает селекцию отдельных групп, видов микроорганизмов и их чрезмерное развитие в почвах. Следствием этого являются стойкие заболевания растений (при развитии патогенных для растений микроорганизмов) (16, 321).
Как правило, в динамике численности микроорганизмов ризопланы и ризосферы наблюдаются два максимума: первый приходится на фазу кущения растений, второй – на фазу цветения и начало плодоношения (см. Приложение 2). В зоне молодого корня доминируют неспорообразующие бактерии рода Pseudomonas и некоторые микроскопические грибы. К фазе цветения растений их сменяют бациллы; актиномицеты, образующие активные вещества – антибиотики, угнетающие развитие патогенов на корнях; клечаткоразрушающие бактерии, которые принимают участие в разложении органических веществ отмирающих корней. Корневые выделения растений, несомненно, служат селективным фактором в формировании микробной ассоциации ризосферы. Например, в ризосфере пшеницы ведущая роль принадлежит микобактериям, в то время как в ризосфере клевера преобладают флюоресцирующие бактерии рода Pseudomonas (26, 82).
Интенсивно протекающие микробиологические процессы трансформации веществ в ризосфере обусловливают накопление в ней водорастворимых элементов минерального питания растений. Выделяемые бактериями угольная и другие минеральные и органические кислоты способствуют растворению и усвоению растениями труднодоступны соединений, таких, как фосфаты кальция, силикаты калия и магния. Синтезируемые микроорганизмами витамины (тиамин, витамин В12, пиридоксин, рибофлавин, пантотеновая кислота и др.) и ростовые вещества (гиббереллин, гетероауксин) оказывают стимулирующее действие на ростовые процессы растений. Многие сапрофитные бактерии ризосферы являются антагонистами фитопатогенных микробов и выполняют роль санитаров в почве.
Ризосферный эффект более ярко выражен в песчаных почвах и менее – в гумусных. В пустынных районах ризосфера является, по-видимому, единственной зоной, где активно развивается микрофлора. В любой почве изменения окружающей среды, включая агротехнические мероприятия, оказывают меньшее воздействие на микроорганизмы в ризосфере по сравнению с обитателями почвы. Ризосферная зона представляет собой своеобразную «буферную» систему, препятствующую воздействию среды на микрофлору (16, 321).
Другой пример симбиоза – сожительство высших растений с клубеньковыми бактериями.
В 1866 г. известный ботаник и почвовед М.С. Воронин увидел в клубеньках на корнях бобовых растений мельчайшие «тельца». Он выдвинул смелые для того времени предположения: связал образование клубеньков с деятельностью бактерий, а усиленное деление клеток ткани корня с реакцией растения на проникшие в корень бактерии (16, 355).
Б. Франк предложил родовое название клубеньковых бактерий – Rhizobium (от греч. Rhizo – корень, bio- жизнь; жизнь на корнях).Это название используется до сих пор. Для обозначения вида клубеньковых бактерий принято к родовому названию добавлять термин, соответствующий латинскому названию того вида растения, на котором они могут образовать клубеньки. Например, Rhizobium trifolii – клубеньковые бактерии клевера.
Для клубеньковых бактерий характерно разнообразие форм: палочковидные, овальные, кокковидные подвижные и неподвижные. Размер колеблется от 0,5-0,9 до 1,2-3 мкм. Делятся перешнуровыванием, а с возрастом могут переходить к почкованию. При старении клубеньковые бактерии теряют подвижность и переходят в состояние опоясанных палочек. Такое название они получили вследствие чередования в клетках плотных и неплотных участков протоплазмы. В двухмесячных клубеньковых бактериях образуется от 1 до 5 артроспор сферической формы. Развиваются при незначительных количествах кислорода в среде, однако, предпочитают аэробные условия. Многие виды клубеньковых бактерий способны синтезировать витамины группы В, а также ростовые вещества типа гетероауксина. Они приблизительно одинаково устойчивы к щелочной реакции среды, но неодинаково чувствительны к кислой.
Для клубеньковых бактерий характерна специфичность – способность образовывать клубеньки. Специфичность может быть узкой: клубеньковые бактерии клевера заражают только группу клеверов – видовая специфичность, а клубеньковые бактерии люпина обладают сортовой специфичностью – заражают только сорта люпина. При широкой специфичности клубеньковые бактерии, например, гороха могут заражать растения гороха, чины, бобов (16, 358).
Процесс внедрения клубеньковых бактерий в ткань корня одинаков у всех видов бобовых растений и состоит из двух фаз.
В первую фазу происходит инфицирование корневых волосков. Проникнув в корень (через корневой волосок, эпидермальную клетку, места повреждений корня), клубеньковые бактерии далее перемещаются в ткани корня растений. Внедриться может одна клетка или группа клеток бактерий, которые передвигаются по корню (16, 362).
Во вторую фазу интенсивно идет процесс образования клубеньков из паренхимных клеток корня и эндодермы (внутреннего слоя первичной коры). В образовании сосудистой системы клубеньков принимают участие ткани центрального цилиндра корня (см. Рис. 4). Защитную функцию у клубеньков выполняет клубеньковая кора, состоящая из нескольких рядов, не зараженных паренхимных клеток корня. Состояние растения-хозяина оказывает влияние на длительность функционирования клубенька.
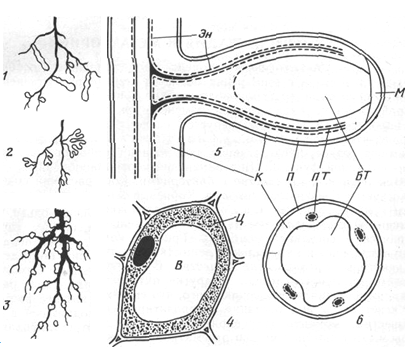
Рис. 4. Бактериальные клубеньки на корнях бобовых:
/— у сочевичника; 2— у клевера ползучего; 3— у люпина; 4— клетка с бактериями в цитоплазме; 5— схема продольного разреза через клубенек и материнский корень; 6—схема поперечного разреза через клубенек; БТ—бактероидная ткань; В—вакуоль; С—кора; М—меристема; Л—перидерма; ПТ—проводящие ткани; Ц—цитоплазма; Эн — эндодерма.
Продолжительность фаз неодинакова у разных видов растений. В некоторых случаях трудно обнаружить границы между фазами. Наиболее интенсивное внедрение клубеньковых бактерий в корневые волоски происходит на ранних этапах развития растения. Вторая фаза заканчивается в период массового образования клубеньков. Нередко внедрение клубеньковых бактерий в корневые волоски продолжается уже после того, как клубеньки сформировались на корнях (16, 362).
В период функционирования клубеньки обычно плотные, образованные активными культурами клубеньковых бактерий и в молодом возрасте имеют беловатую окраску. К моменту проявления оптимальной активности они становятся розовыми. Клубеньки, возникшие при заражении (инфекции) неактивными культурами бактерий, зеленоватого тона. Старые образования тёмные, дряблые, мягкие. При надрезе из них выступает водянистая слизь. Клубеньки многолетних растений, в отличие от клубеньков однолетних, могут функционировать в течение многих лет. К концу вегетационного периода бактероидная ткань многолетних клубеньков деградирует, но всё образование не отмирает. На следующий год клубенёк вновь начинает функционировать.
Для симбиоза, обеспечивающего хорошее развитие растений, необходим определенный комплекс условий среды: влажность, температура, реакция почвы, доступ кислорода, наличие определенных химических элементов. Большое значение имеет влияниие ризосферной микрофлоры на клубеньковые бактерии, которое могут быть стимуляционным или антагонистическим в зависимости от состава микроорганизмов ризосферы (16, 373).
Из 13.000 видов бобовых растений наличие клубеньков выявлено пока только приблизительно у 1.300 видов. Сюда в первую очередь относятся виды растений, использующиеся в сельском хозяйстве - более 200. Сформировав клубеньки, бобовые растения приобретают способность усваивать атмосферный азот, который фиксируют из воздуха. Азотфиксирующие бактерии восстанавливают азот (N2) до аммиака (NH3) с помощью сложной ферментной системы нитрогеназы содержащей железо, молибден, магний. Эта система нуждается в источнике электронов, которые поступают в нитрогеназную систему через восстановитель с низким потенциалом, содержащий негеминовое железо – ферродоксин (переносчик электронов) (см. Рис. 5).
 Донор Фд·2ē (АзоФд)2·2ē МоФд·2ē
Донор Фд·2ē (АзоФд)2·2ē МоФд·2ē
![]()
![]()
![]()
![]()
![]() электронов / / / АТФ АДФ+2NH3 +Ф
электронов / / / АТФ АДФ+2NH3 +Ф
![]()
![]()
![]() Фд / (АзоФд)2 / МоФд /
Фд / (АзоФд)2 / МоФд /
Рис. 5. Восстановление азота до аммиака азотфиксирующими бактериями.
Цепь переноса электронов состоит из ферредоксина (Фд), азоферредоксина (АзоФд) и молибдоферредоксина (МоФд) и за один раз переносятся только два электрона. Для последнего переноса расходуется одна молекула АТФ (21, 53).
Растения поставляют клубеньковым бактериям продукты углеводного обмена и минеральные соли, необходимые им для роста и развития.
Часто встречается микоризообразование – симбиоз мицелия гриба с корнем высшего растения. Это явление открыл в 1881 г. польский ученый Ф.М. Каменский, которое в последствии получило название микоризы (от греч. mykes – гриб и rhiza – корень, грибокорень) (9, 142).
При этом в непосредственный контакт вступает грибница, находящаяся в почве. По тому, как осуществляется этот контакт, различают два типа микоризы – эндотрофную и экзотрофную (17, 232). У эндотрофных микориз мицелий гриба распространен главным образом внутри тканей корня (в коровой паренхиме) и относительно немного выходит наружу (см. Рис. 6). Корни при этом несут нормальные корневые волоски. Мицелий гриба идет и межклетно, и внутриклетно. Типичная эндотрофная микориза характерна для семейства (Orchidaceae), где она также является для большинства видов облигатной, то есть семена этих растений не могут прорастать и развиваться в отсутствие гриба. Эндотрофная микориза встречается у многих травянистых растений, хотя присутствие гриба для развития растения здесь не столь обязательно, реже – у деревьев.
Эктотрофная микориза отличается наличием наружно чехла из гиф на корнях, или она имеет вид плотно переплетенной ткани, одевающей корень (см Рис. 6). От этого переплетения гиф в окружающую почву простираются свободные гифы. Собственных корневых волосков корень при этом не образует.
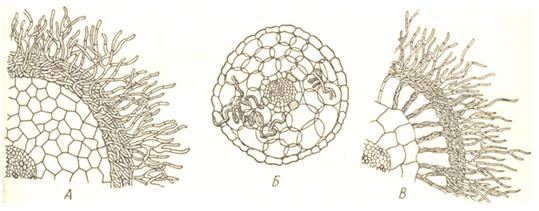
Рис. 6. Поперечные срезы микориз древесных пород. Микоризы: А – эктотрофная (береза); Б – эндотрофная (клен); В – эктоэндотрофная (дуб).
Гифы гриба проходят между клетками эпидермиса, образуя сеть. Такая микориза характерна для древесных растений и редко встречается у травянистых (4, 221).
Провести четкую границу между эндотрофной и экзотрофной мико-ризами трудно. Переходом между ними являются эктоэндотрофная микориза, распространенная более часто, чем чисто эктотрофная. Грибные гифы при такой микоризе густо оплетают корень снаружи и в то же время дают обильные ветви, проникающие в его коровую паренхиму. Мицелий идет отчасти межклетно, отчасти внутриклетно, образуя в клетках клубки, гиф или разветвления, напоминающие гаустории. Клетки корня при этом остаются живыми (см. Рис.6). Такая микориза встречается у большинства древесных пород: дуб, ель, сосна, лиственница и другие (17, 233).
Микоризу образует большинство растений (за исключением водных), как древесных, так и травянистых (особенно многолетних). Травянистые растения вступают в микоризный симбиоз с микроскопическими грибами в основном из класса несовершенных грибов (Deuteromycetes), отчасти из класса зигомицетов (Zygomycetes) с мицелием, лишенным перегородок (неклеточным) и отчасти из класса сумчатых грибов (Ascomycetes). Грибы родов элафомицес (Elaphomyces) и трюфель (Tuber) образуют микоризу с буком, дубом и другими деревьями. Но большинство древесных пород образуют микоризу с грибницей шляпочных грибов – макромицетов из класса базидиальных (Basidiomycetes) и группы порядков гименомицетов.
В гименомицетной эктоэндотрофной микоризе древесных пород гриб оплетает корень снаружи и частично проникает внутрь. Здесь он получает от корня углеводное питание, так как сам, будучи гетеротрофом, не может синтезировать органические вещества. Наружные свободные гифы гриба широко расходятся в почве от корня, заменяя последнему корневые волоски. Эти свободные гифы получают из почвы воду, минеральные соли, а также растворимые органические вещества (главным образом азотистые). Часть этих веществ поступает в корень, а часть используется самим грибом на построение грибницы и плодовых тел. В клетках корня частично перевариваются внедрившиеся гифы и служат дополнительным источником органических веществ.
Почва леса, особенно в прикорневой зоне деревьев, пронизана грибницей микоризных грибов, а на поверхности почвы появляются многочисленные плодовые тела. Это подберезовик (Leccinum scabrum), подосиновик (Leccinum aurantiacum) и многие другие шляпочные грибы, встречающиеся только в лесу. Для этих грибов такой симбиоз обязателен. Если их грибница и может развиваться без участия корней дерева, то плодовые тела в этом случае обычно не образуются.
Белый гриб образует микоризу со многими породами деревьев: березой, дубом, сосной, елью. Некоторые виды грибов – только с одной определенной породой. Так лиственничный масленок образует микоризу с лиственницей.
Для деревьев симбиоз с грибами тоже имеет значение: опыты на лесных полосах показали, что без микоризы деревья развиваются хуже, отстают в росте, они более ослаблены, подвержены заболеваниям.
В случае эндотрофных микориз взаимоотношения гриба и высшего растения еще более сложные. В связи с малым контактом гиф микоризного гриба с почвой таким путем в корень поступает относительно небольшое количество воды, минеральных и азотистых веществ. В этом случае значение для высшего растения, вероятно, приобретают вырабатываемые грибом биологически активные вещества типа витаминов. Отчасти гриб снабжает высшее растение азотистыми веществами, так как часть гиф гриба, находящаяся в клетках корня, переваривается ими. Гриб получает углеводы, а в случае микоризы орхидных гриб сам отдает углеводы (сахар) высшему растению (17, 234).
Нужно отметить, что микоризный симбиоз – явление очень сложное, не укладывающееся в какую-то определенную схему. Эти взаимоотношения часто зависят от окружающих условий.
Некоторые микроорганизмы ризосферы постепенно переходят в состав эпифитной микрофлоры. Особенно оживился этот вопрос в связи с работами немецкого ученого Е. Либберта (1966), который выступил с утверждением, что гормон роста высших растений гетероауксин синтезируется не растением, а эпифитной микрофлорой.
Работы В.И. Кефели (1969, 1971) показали, что в стерильных условия капуста образует гетероауксин из L – триптофана. А.А. Тарасенко (1972) установила, что эпифитная микрофлора влияет положительно на рост и обмен веществ проростков кукурузы (19, 142).
Микроорганизмы, развивающиеся на поверхности стеблей или листьев растений, получили название эпифитной микрофлоры. Микроорганизмы-эпифиты вынуждены довольствоваться минимальными источниками питательного субстрата, представленного выделениями растительных тканей и веществами-загрязнителями (пылью). Поэтому состав эпифитной микрофлоры весьма специфичен. Нередко 80% общего количества эпифитов составляют бактерии Erwinia herbicola. Второе место по численности занимают различные грибы (Penicillum, Mucor, Fusarium и другие). На поверхности многих тропических растений обнаружены азотфиксирующие бактерии рода Beiyerckia, поставляющие азот непосредственно в лист.
Разнообразная и обильная микрофлора находится на поверхности семян. Так, на 1г зерна ржи приходится не менее 2.500 тыс. микробных клеток, пшеницы – 1.500 тыс., риса – 250 тыс. В состав микрофлоры зерна обязательно входят неспорообразующие бактерии Pseudomonas, Arthrobacter и Flavobacterium, дрожжи Candida, Rhodotorula, Criptococcus, а также грибы Penicilium, Aspergillis, Alternaria, Cladosporium, Mucor и другие. Развитие микроорганизмов на поверхности зерна в значительной мере зависит от влажности и температуры. Установлено, что при температуре 15 – 20°С и влажности 14,5 – 15% на зерне пшеницы начинается развитие грибов, а при влажности 17,5 – 18% - бактерий.
Общая численность эпифитных микроорганизмов резко возрастает при повышении влажности воздуха и усиленном выделении продуктов обмена растительными тканями.
При жизни растения эпифитная микрофлора не оказывает на него вредного влияния. Наоборот, питаясь продуктами выделений растений, она способствует освобождению тканей от продуктов собственного обмена (15, 259). Эпифитная микрофлора образует определенный биологический барьер, препятствующий заражению растительных тканей фитопатогенными микробами (26, 83).
Таким образом, симбиотические взаимоотношения микроорганизмов с высшими растениями разнообразны. При таком контакте пользу получают оба организма, не нанося друг другу вреда.
Похожие работы
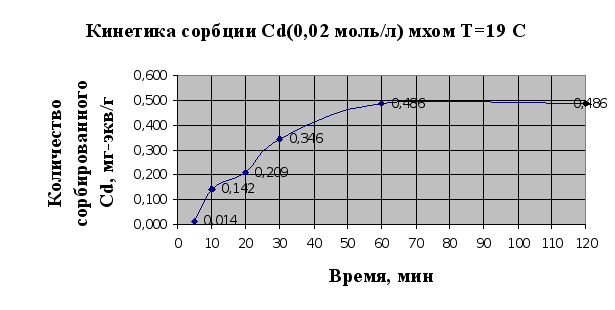
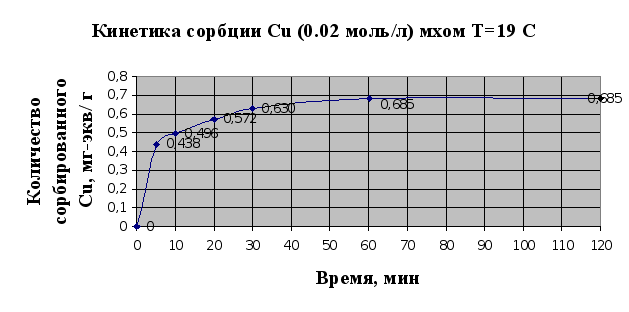
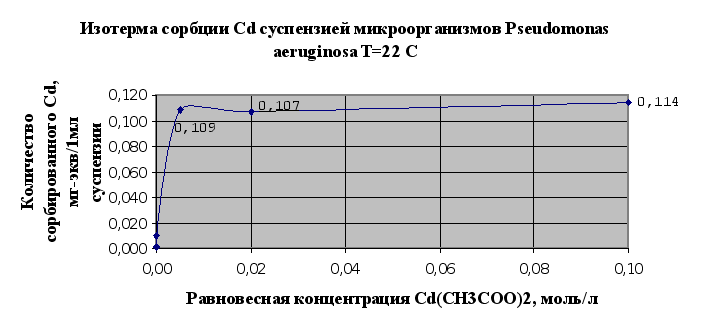
... Листов Пров. 1 2 Консульт. БГТУ 7140607 2004 Н. контр. Утв. Целью данной дипломной работы является изучение сорбционных свойств мха по отношению к микроорганизмам и тяжелым металлам, а также изучение сорбционной способности системы «мох-микроорганизмы» и выявление доли участия каждого компонента этой ...
... рассмотренный в тексте обзора межклеточный матрикс. Настоящая работа имеет и ещё один аспект. Рассмотренные в ней данные последних десятилетий показывают, что адекватно понять колониальную организацию и межклеточную коммуникацию микроорганизмов можно лишь в том случае, если учесть всю гамму не только внутривидовых, но и межвидовых экологических отношений. Иначе, говоря биосоциальные микробные ...
... два других отдела, отделенных мембраной, пирреллюлозому или рибоплазму, которая содержит рибосомы и связанные белки, и свободный от рибосом парафоплазму (Glockner, 2003). 3. Характеристика общих свойств микроорганизмов Микроорганизмы - это организмы, невидимые невооруженным глазом из-за их незначительных размеров. Этот критерий - единственный, который их объединяет. В остальном мир ...
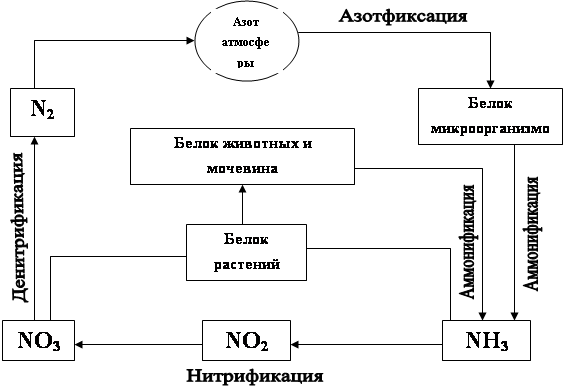
... в какой последовательности и как глубоко учащиеся будут изучать материал. По программе Сивоглазова В.И., Сухова Т.С., Козлова Т.А. в книге для учителя «Биология: общие закономерности» тема «Биогеохимическая деятельность микроорганизмов» не рассматривается как самостоятельная на отдельном уроке, а является составной частью других тем. Например, на уроке по теме «Значение прокариот в биоценозах, их ...
0 комментариев