Навигация
Транспорт липидов у эукариот
48348
знаков
0
таблиц
5
изображений
3.3.2 Транспорт липидов у эукариот
Механизм распределения фосфолипидов в эукариотических клетках гораздо более сложен. Единственным фосфолипидом, который локализуется исключительно в месте своего синтеза, в митохондрии, является кардиолипин. Как показывают многочисленные эксперименты, внутриклеточный транспорт фосфолипидов осуществляется значительно быстрее, чем если бы он определялся самопроизвольной диффузией в водной среде.
Например, перенос новосинтезированного фосфолипида от эндоплазматического рети-кулума к митохондриям в клетках печени крысы происходит всего за несколько минут. Поскольку декарбоксилаза, катализирующая превращение фосфатидилсерина в фосфатидилэтаноламин, содержится в митохондриях, данные о скорости этой реакции могут использоваться для контроля за переносом фосфатидилсерина из эндоплазматического ретикулума в митохондрию. Показано, что скорость переноса весьма высока и процесс ингибируется азидом и агентами, блокирующими биоэнергетические реакции.
Обнаружено также, что новосинтезированный фосфолипид в течение нескольких минут транспортируется к плазматической мембране, хотя скорость переноса очень сильно зависит от самого липида и типа клетки. В некоторых случаях транспорт липидов блокируется ингибиторами биоэнергетических реакций (энергетическими ядами) и/или агентами, разрушающими цитоскелет. Общепринято, что скорость переноса липидов выше скорости диффузии.
Одним из способов, позволяющих облегчить перенос фосфолипидов между мембранами, является участие в этом процессе белков. Выделен целый ряд водорастворимых белков, участвующих в межмембранном переносе фосфолипидов in vitro. Все они имеют липидсвязывающие участки и образуют водорастворимые комплексы с фосфолипидами. Эти белки облегчают обмен липидов «один к одному» между мембранами путем переноса мономеров.
В одних случаях (например, для фосфатидилхолинспецифического белка переноса из печени быка) связывание липндов происходит с высокой специфичностью и сродством, в других сродство и специфичность относительно низки. В принципе при низком сродстве способность белка катализировать перенос липидов от одной мембраны к другой должна повышаться, поскольку белок при этом может возвращаться к донорной мембране, имея незанятый липидсвязывающнй участок. Белки, переносящие фосфолипиды, были выделены не только из животных клеток, но и из бактерий и клеток растений. Очищены белки, связывающиеся сгликолипидами и длинноцепочечными жирными кислотами,
К сожалению, функция белков, переносящих фосфолипиды, до сих пор не продемонстрирована in vivo, поэтому неясно, играют ли они физиологическую роль во внутриклеточном переносе фосфолнпидов. Не исключено, что этот перенос опосредуется везикулами, а возможно, функционируют оба механизма.
Важно понять, что так или иначе распределение фосфолипидов среди различных клеточных мембран детерминируется белками. Если распределение липидов равновесно, то оно определяется сродством белков, присутствующих в разных мембранах, к липидам. Смещение в распределении некоторых липидов из-за взаимодействия их с белками может влиять на распределение других липидов, например холестерола . Если распределение липидов неравновесно, то липидный состав разных мембран будет определяться скоростью транспортировки липидов к мембранам и от них, но и в этом случае кинетика процесса будет зависеть от участия в нем определенных белков.
3.4 Изменение состава липидов под действием условий внешней среды
Из всего сказанного ясно, что о механизме контроля липидного состава клеточных мембран известно очень мало. Одно из минимальных требований состоит в том, что мембранные липиды должны образовывать стабильный бислой, находящийся в жидкокристаллическом состоянии. Механизмы, обеспечивающие изменение липидного состава мембран в ответ на изменения условий окружающей среды, имеют многие организмы. Лучше всего изучен механизм тепловой адаптации Е. coli.
1. Тепловая адаптация Е. Coli.. При выращивании Е. coli в условиях низких температур наблюдается изменение жирнокислотного состава в сторону большего содержания ненасыщенных жирных кислот. Это способствует поддержанию мембраны в текучем состоянии и позволяет клеткам выжить при экстремальных температурах. Преобладающими остатками жирных кислот в Е. coli) являются пальмитоил (16:0), пальмитолеил (16:1) и цис-ваксеноил (18:1). При низких температурах в бислой включается больше дос-ваксеновой кислоты, а содержание пальмитолеиловой кислоты остается постоянным. Связано это с функционированием одного из двух ферментов, которые катализируют удлинение жнр-нокислотных цепей. При низких температурах фермент 3-кетоацил-АСР-синтаза II более активно превращает пальмитолеиловую кислоту в дос-ваксеновую, увеличивая пул ненасыщенных жнриых кислот, включаемых в фосфолипиды.
2. Адаптация к повышению давления. Жирнокнслотный состав барофильной морской бактерии NPT3 зависит от давления. При изменении давления в мембранах может также происходить фазовый переход, и эта адаптация, как и в предыдущем случае, позволяет организму выжить. При повышении давления содержание в мембранах ненасыщенных жирных кислот увеличивается, и благодаря особеиностям их упаковки мембрана остается в жидкокристаллическом состоянии. Аналогичные результаты были получены при изучении фосфолипидов митохондрий из клеток печени глубоководных океанических рыб.
4 Заключение
Клетки эукариот содержат много мембранных органелл и множество различных внутриклеточных мембран, каждая из которых обладает уникальным белковым и липидным составом. Любой мембранный белок, информация о синтезе которого заключена в ядре, должен безошибочно доставляться от места синтеза на риоо-соме, находящейся в цитоплазме, к месту назначения. Для этого используется сложная система сигнальных последовательностей, содержащихся в любой зрелой форме полипептида или предшественника, а также рецепторы внутри клетки, способные эти сигналы распознавать. Некоторые мембранные белки включаются в липидный бислой самопроизвольно, но в большинстве случаев правильная сборка белка внутри клеточной мембраны является энергозависимым процессом, который осуществляется с помощью специализированного аппарата. По-видимому, белки не могут включиться в клеточную мембрану до тех пор, пока они не приобретут частично развернутую конформацию. Разворачивание белков или поддержание их в развернутой конформацин, необходимой для переноса, возможно, осуществляются при участии АТР и специфических белков в цитоплазме.
В эукариотической клетке большинство липидов синтезируется в эндоплазматическом ретикулуме, и чтобы попасть к месту назначения, они должны пройти через соответствующие мембраны. Механизм транспорта липидов неизвестен, можно лишь предположить, что в нем участвуют везикулы и белки, связывающие липиды. Способ поддержания липидного состава различных внутриклеточных мембран тоже не установлен.Литература
1.Р. Геннис «Биомембраны, молекулярная структура и функции» Москва «Мир» 1997 г.
2.В.К. Рубальченко «Структура и функции мембран» Киев 1988г.
3. Болдырев А.А. Лекции по биохимии мембран.
4. Финдлей Дж. Биохимия мембран. Методы. «Мир » 1990г.
Похожие работы
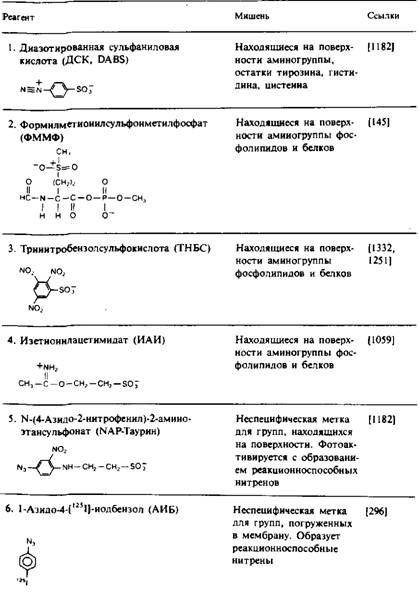

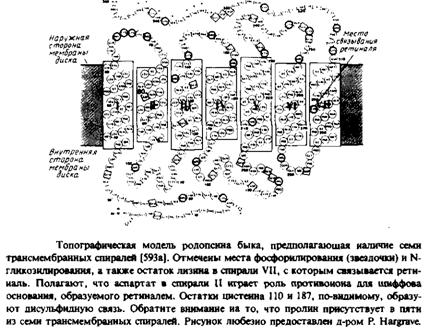


... и индуцируют трансмембранную миграцию липидных молекул, поэтому полученные результаты бывает трудно интерпретировать. Детальная оценка достоинств и недостатков методов изучения липидной асимметрии в мембранах дается, например, в обзорах. МЕТОДЫ УСТАНОВЛЕНИЯ ТРАНСМЕМБРАННОГО РАСПРЕДЕЛЕНИЯ ЛИПИДОВ Химическая модификация фосфолипидов Относительно легко подвергаются химической модификации только ...
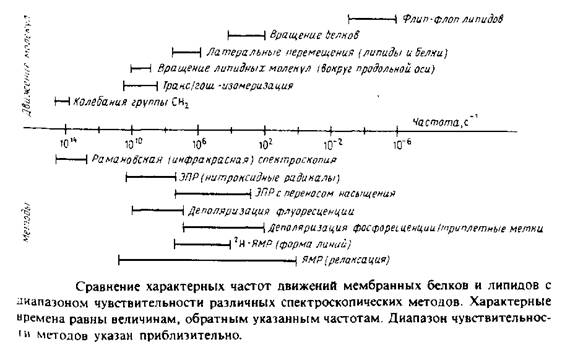
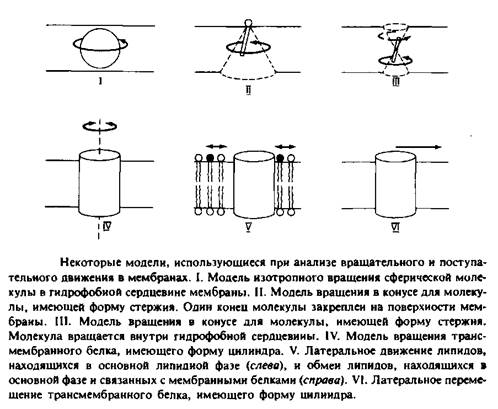
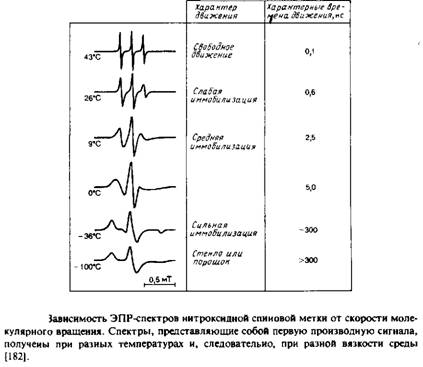
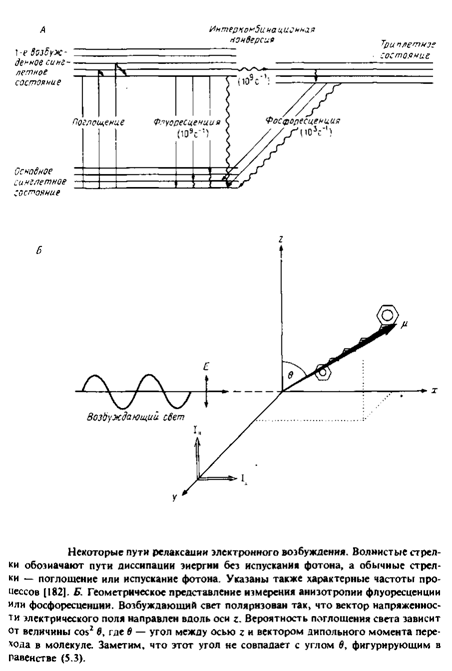
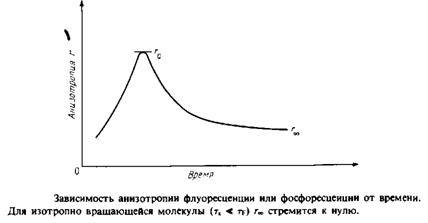
... к уменьшению латеральной диффузии. 5. Липидно-белковые взаимодействия Большинство методов, применяемых для изучения упорядоченности и динамических свойств мембран, используется и для исследования липидно-белковых взаимодействий. Работы по изучению этих взаимодействий были в основном направлены на выяснение влияния мембранных белков на физическое состояние липидов. Рассмотрим типичную мембрану ...
... является курсом, для изучения которого необходимо наличие знаний об основных принципах организации биологических молекул, строении и механизмах действия ферментов. Дисциплина биохимия мембран относится к дисциплинам специализации федерального компонента. 5. Распределение времени, отведенного на изучение дисциплины по учебному плану Форма учебной работы Форма обучения Очная По ...
... линейной последовательности нуклеотидов) явилось качественным переходом химической эволюции на свою следующую ступень или отправной точкой для самой ранней стадии биогенеза, непосредственно предшествующей процессу возникновения жизни. Следовательно, можно с определённой долей уверенности предположить, что стадия накопления „правильного“ для биогенеза химического потенциала сумела подготовить ...
0 комментариев