Навигация
СРАВНЕНИЕ ХОРДОВЫХ С ДРУГИМИ ВТОРИЧНОРОТЫМИ
29611
знаков
1
таблица
8
изображений
4. СРАВНЕНИЕ ХОРДОВЫХ С ДРУГИМИ ВТОРИЧНОРОТЫМИ
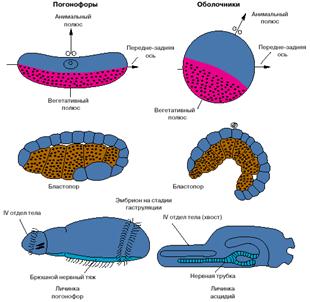
Хотя хордовые - это вторичноротые животные, все же их развитие имеет ряд особенностей. На рис. 4 представлены соответствующие стадии развития одной из групп вторичноротых - погонофор и низших хордовых - асцидий.
Анимальный полюс яйца погонофор, на котором располагается ядро яйцеклетки, соответствует спинной стороне эмбриона, а вегетативный, на котором концентрируется желток, - брюшной. У низших хордовый ситуация обратная: анимальный полюс соответствует брюшной стороне, а вегетативный - спинной. Бластопор (то есть та область эмбриона, где впячивается энтодерма и формируется первичный кишечник) у погонофор соответствует брюшной стороне личинки, а у низших хордовых - спинной. Зачаток нервной системы, закладывающийся по линии замыкания бластопора, у погонофор становится брюшным нервным тяжем, а у хордовых - спинной нервной трубкой (рис. 4).
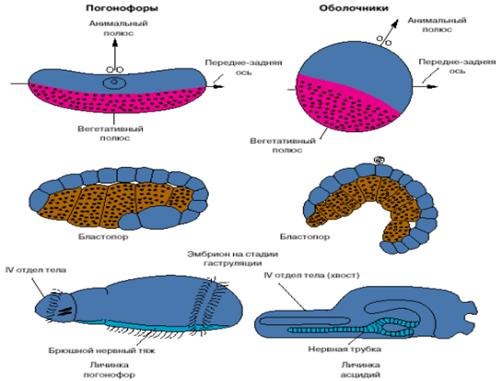
Рис. 4. Особенности эмбрионального развития погонофор и низших хордовых.
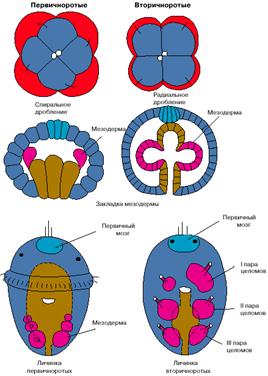
План строения низших хордовых также существенно отличается от планов строения других вторичноротых. Так, например, у всех вторичноротых передние целомы диссимметричны: левый передний целом больше правого, который обычно выполняет функцию околосердечной сумки. У хордовых ситуация обратная: правый передний целом больше левого. Целомы вторичноротых сообщаются с внешней средой отверстиями, открывающимися на спинной стороне и выполняющими осморегуляторные и половые функции. У хордовых эти отверстия располагаются на той стороне тела, которую мы традиционно считаем брюшной.
Кровеносная система у вторичноротых и хордовых устроена по общему плану и состоит из двух главных сосудов: спинного и брюшного. У вторичноротых, как и всех других беспозвоночных, кровь по спинному сосуду течет вперед, а по брюшному назад. У хордовых направление тока крови обратное: по спинному сосуду кровь течет назад, а по брюшному - вперед. Сердце у вторичноротых лежит на спинной стороне, а у хордовых - на брюшной.
Суммируем все эти отличия.
1. Бластопор хордовых соответствует не брюшной, а спинной стороне.
2. Нервная пластинка хордовых, закладывающаяся по месту замыкания бластопора, превращается не в брюшной нервный тяж, а в спинную нервную трубку.
3. Диссимметрия первой пары целомов имеет обратный знак в сравнении с другими вторичноротыми: правый передний целом у хордовых больше левого.
4. Отверстия целомов у хордовых открываются не на спинной, а на брюшной стороне.
5. Кровь у хордовых по брюшной стороне течет вперед, а по спинной - назад, и сердце располагается не на спинной, а на брюшной стороне.
Это значит, что предки хордовых в своем эволюционном развитии испытали инверсию сторон тела, то есть перевернулись и стали передвигаться на морфологически спинной стороне, которая стала функционировать как физиологически брюшная. Морфологически брюшная сторона предков хордовых стала функционировать как физиологически спинная.
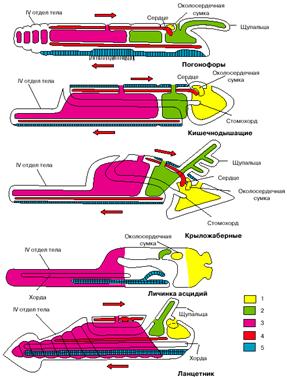
Если мы примем этот вывод, все перечисленные выше несоответствия исчезнут: бластопор хордовых станет соответствовать их исходно брюшной стороне, нервная трубка будет располагаться на морфологически брюшной стороне, диссимметрия передних целомов приобретет нормальный для вторичноротых характер, отверстия целомов окажутся на морфологически спинной стороне, и направление тока крови, так же как положение сердца, окажется типичным для вторичноротых. На рис. 5 представители низших хордовых (личинка асцидий и ланцетник) изображены перевернутыми, чтобы продемонстрировать, что при таком положении их системы органов соответствуют таковым других вторичноротых.
Переворот на спинную сторону мог стать тем важнейшим эволюционным событием, которое определило формирование плана строения хордовых животных и его коренное отличие от планов строения вторичноротых и других беспозвоночных.
5. ПУТИ ЭВОЛЮЦИИ ВТОРИЧНОРОТЫХ ЖИВОТНЫХ
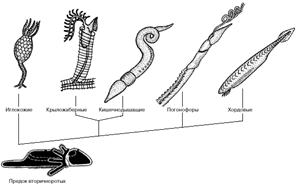
Общий предок вторичноротых животных, по-видимому, вел донный образ жизни, ползая на брюшной стороне с помощью ресничной полоски, подостланной вентральным нервным тяжем (рис. 6). Он имел три пары целомических мешков и щупальцевый аппарат, с помощью которого собирал мелкие пищевые частицы. В дальнейшем могло произойти обособление самого заднего отдела тела, с помощью которого предки вторичноротых временно зарывались в верхние слои грунта. В эволюции вторичноротых этому отделу принадлежит очень важное место. Предки иглокожих перешли к сидячему образу жизни и стали собирать взвешенные в воде пищевые частицы с помощью щупалец. Радиальная симметрия иглокожих - это наследие сидячего образа жизни, который среди современных форм сохраняют морские лилии, тогда как морские звезды, морские ежи, офиуры и голотурии способны медленно переползать с места на место.
Полухордовые в качестве основного органа движения стали использовать предротовой отдел тела: хобот кишечнодышащих или головной щит крыложаберных. Обычно полухордовых считают близкими предками хордовых, о чем говорит и само название этой группы.
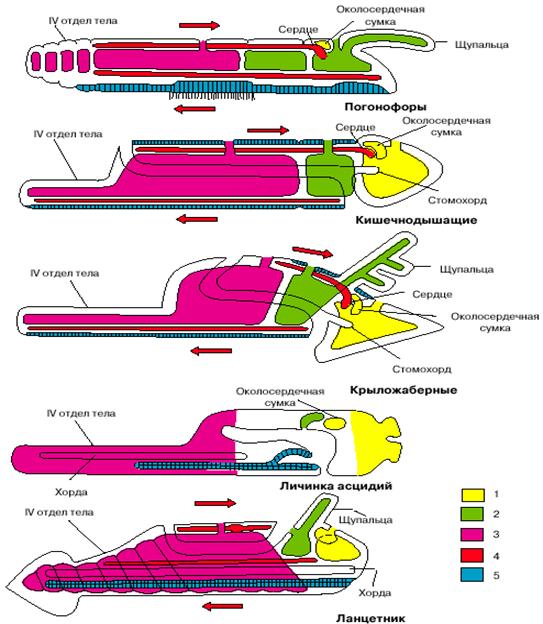
Рис. 5. Сравнение планов строения различных групп вторичноротых (личинка асцидий и ланцетник изображены в перевернутом положении). 1 - целомы первой пары, 2 - целомы второй пары, 3 - целомы третьей пары, 4 - кровеносная система, 5 - нервная система. Стрелки показывают направление тока крови.
В действительности эта группа не слишком близка к хордовым, в частности потому, что имеет два нервных тяжа, только один из которых может быть гомологизирован с нервной трубкой хордовых. У них имеется слепой вырост кишечника - стомохорд, который сформировался как опорная структура хобота и не имеет прямого отношения к хорде хордовых животных.
Современные погонофоры - сидячие организмы, обитающие внутри выделяемых ими трубок. Во взрослом состоянии они лишены кишечника и питаются за счет симбиотических хемосинтезирующих бактерий, окисляющих сероводород или метан. Их предки, вероятно, питались этими бактериями, собирая их из придонного слоя воды с помощью щупалец.
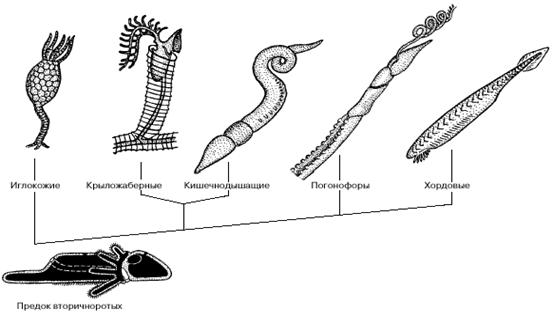
Рис. 6. Филогенетические отношения между вторичноротыми животными.
Впоследствии они стали культивировать бактерий в клетках кишечника. Личинки одной из групп современных погонофор - вестиментифер - на ранних стадиях развития собирают бактерий с поверхности грунта, а затем начинают их культивировать внутри видоизмененного кишечника, после чего рот и анус у них редуцируются. Особенности эмбрионального развития и строения нервной системы позволяют рассматривать погонофор как самую близкую к хордовым группу вторичноротых.
Предки хордовых также первоначально собирали пищевые частицы с помощью околоротовых щупалец, которые до сих пор сохранились у ланцетников. Одной из особенностей эволюции хордовых явилось использование самого заднего, четвертого отдела тела как органа активного движения. Этот отдел имеется у всех вторичноротых. Иглокожие и крыложаберные используют его как прикрепительный стебелек. У кишечнодышащих этот отдел функционирует только у молодых особей: с его помощью они закапываются в толщу грунта. У погонофор этот отдел активно функционирует всю жизнь: он вооружен щетинками, торчит из заднего конца трубки и благодаря его движениям погонофоры заглубляют задний конец трубки в толщу грунта по мере своего роста. Хордовые - единственная группа вторичноротых, использующая этот отдел как орган активного плавания. Именно в связи с активной двигательной функцией этого органа в нем происходит полная сегментация мускулатуры и развивается хорда (у личинок оболочников хорда локализована именно в хвосте. Другой важный момент эволюции хордовых - это инверсия сторон тела, то есть произошедший в филогенетическом развитии хордовых переворот с морфологически брюшной на морфологически спинную сторону. Такой переворот мог совершиться при переходе от улавливания с помощью щупальцевого аппарата взвешенных в толще воды пищевых частиц, как это делают, например, современные крыложаберные, к их сбору из поверхностного слоя грунта, как это делают современные ланцетники.
Заметим, что между личинками оболочников, с одной стороны, и бесчерепными и позвоночными, с другой, есть одно важное различие. Если внимательно посмотреть на две нижние фигуры на рис. 5, изображающие личинку асцидии и ланцетника, то нетрудно заметить, что у первых рот располагается на той же стороне, что и нервная трубка (последняя даже открывается в глотку особым отверстием), то есть на исходно брюшной стороне, а у ланцетника (так же, как у позвоночных) - на противоположной стороне тела. Положение рта у личинок асцидий - первичное, оно совпадает с тем, что имеет место у других вторичноротых (см. рис. 4). Таким образом, личинки асцидий по своей организации более примитивны, чем остальные хордовые, и отражают организацию подвижных предков хордовых до их переворота.
У бесчерепных и позвоночных рот располагается на противоположной стороне, гомологичной спинной стороне тела прочих вторичноротых (см. рис. 5). Такое положение рта не может быть первичным и возникло уже после переворота. Удивительно, но в индивидуальном развитии бесчерепных сохранились следы постепенного перемещения рта со спинной стороны на брюшную по левой стороне тела. А один из представителей бесчерепных (Assymetron) так и застыл на стадии, когда рот остался на левой стороне тела. Все это отголоски постепенного перемещения рта со старой морфологически брюшной стороны на новую физиологически брюшную сторону.
Похожие работы
... . ЛИТЕРАТУРА: 1. Курс низших растений //Под ред. М. В. Горленко – М.: Высш. школа, 1981. 2. Жизнь растений, т. 1–3, - М.: Просвещение, 1974-77. 3. Н. Грин, У. Стаут, Д. Тейлор. Биология, т. 1-3. – М.: Мир, 1990. 4. Н.П.Горбунова , Е.С.Клюшникова, Н.А.Комарницкий. и др. Малый практикум по низшим растениям. – М.: Высш. школа, 1976. 5. Русанов В. А. Низшие растения. Методические указания ...
... . Переворот на спинную сторону мог стать тем важнейшим эволюционным событием, которое определило формирование плана строения хордовых животных и его коренное отличие от планов строения вторичноротых и других беспозвоночных. Происхождение хордовых животных. В. В. МАЛАХОВ. Продолжение. Пути эволюции вторичноротых животных Общий предок вторичноротых животных, по-видимому, вел донный образ жизни ...

... , которые, кроме семян хвойных, едят много семян злаков и бобовых, мыши, которые, в отличие от полевок, едят сравнительно мало травы. Семеноеды сравнительно ограничены в добыче пищи, и успех их жизнедеятельности зависит зачастую от урожая семян немногих видов растений. Неурожаи таких кормов влекут за собой массовые миграции зверей или их гибель. Так, например, наша белка в годы неурожая хвойных ...
... . Утрата личиночных стадий и/или стадий бесполого размножения: пресноводные гидры, олигохеты, большинство брюхоногих моллюсков. Утрата конечных стадий и размножение на ранних этапах онтогенеза: неотения. Таким образом, существует множество основных типов онтогенеза и еще большее число производных типов. В теории эволюции обычно рассматривается онтогенез на примере цветковых растений и позвоночных ...
0 комментариев