Навигация
2.2. Стохастические модели
Детерминистические модели позволяют эффективно описывать динамику распределения генов в эволюционирующих популяциях. Однако эти модели основаны на предположении бесконечного размера популяции, которое является слишком сильным для многих реальных случаев. Чтобы преодолеть это ограничение, были разработаны вероятностные методы теоретической популяционной генетики [1,3,4,6-8]. Эти методы включают анализ с помощью цепей Маркова (в частности, метод производящих функций) [4,7], и диффузионные [1,3,4,6,8] методы.
Ниже мы кратко рассмотрим основные уравнения и характерные примеры применения диффузионного метода. Этот метод достаточно нетривиален и его применение приводит к достаточно содержательным результатам.
2.2.1. Прямое и обратное уравнения Колмогорова
Рассмотрим популяцию диплоидных организмов с двумя аллелями A1 и A2 в некотором локусе. Численность популяции n предполагается конечной, но достаточно большой, так что частоты гена могут быть описаны непрерывными величинами. Мы также предполагаем, что численность популяции n постоянна.
Введем функцию j = j (X,t|P,0) , которая характеризует плотность вероятности того, что частота гена A1 равна X в момент времени t при условии, что начальная частота (в момент t = 0) была равна P. В предположении малого изменения частот генов за одно поколение, динамика популяции может быть описана приближенно следующими дифференциальными уравнениями в частных производных [1,3,4,8]:
¶ j /¶ t = - ¶ (MdX j )/¶ X + (1/2)¶ 2(VdX j)/¶ X 2 , (6)
¶ j/¶ t = Md P ¶ j/¶ P + (1/2)VdP ¶ 2j/¶ P 2 , (7)
где MdX , Md P и VdX , VdP – средние значения и дисперсии изменения частот X, P за одно поколение, соответственно; единица времени равна длительности одного поколения. Уравнение (6) есть прямое уравнение Колмогорова. (В физике это уравнение называют уравнением Фоккера-Планка), уравнение (7) – обратное уравнение Колмогорова.
Первые слагаемые справа в уравнениях (6), (7) описывают давление отбора, которое обусловлено разностью приспособленностей генов A1 и A2. Вторые слагаемые характеризуют случайный дрейф частот, который обусловлен флуктуациями в популяции конечной численности.
Используя уравнение (6), можно определять динамику частот генов во времени. Уравнение (7) позволяет оценивать вероятности фиксации генов.
Предполагая, что 1) приспособленности генов A1 и A2 равны 1 и 1 - s , соответственно и 2) вклады генов в приспособленности генных пар A1 A1, A1 A2 и A2 A2 аддитивны, можно получить, что величины MdX , Md P и VdX , VdP определяются следующими выражениями [1,3,4,8]:
Md X = sX(1-X), Md P = sP(1-P), Vd X = X(1-X)/(2n), VdP = P(1-P)/(2n) . (8)
2.2.2. Случай чисто нейтральной эволюции
Если эволюция чисто нейтральная (s = 0), то уравнение (6) принимает вид:
¶ j/¶ t = (1/4n)¶ 2[X(1-X)j]/¶ X 2 . (9)
Это уравнение было решено аналитически М. Кимурой [1,6]. Само решение имеет сложный вид, основные результаты этого решения сводятся к следующему: 1) в конечной популяции фиксируется только один ген (A1 либо A2); 2) типичное время перехода от начального распределения к конечному составляет величину порядка 2n поколений. Отметим, что этот результат согласуется с оценками лекции 4 , где была рассмотрена несколько иная модель "чисто нейтральной" эволюции.
2.2.3. Вероятность фиксации гена
Используя уравнение (7), мы можем оценить вероятность фиксации гена A1 в конечной популяции. Действительно, рассматривая асимптотику при времени, стремящемся к бесконечности ( t --> inf ), мы можем положить ¶ j /¶ t = 0 и X = 1 ; тогда аппроксимируя вероятность u(P) , которую нужно найти, величиной u(P) = j (1, inf |P,0)/(2n) (здесь u(P) = j(1, inf |P,0)DX , где DX = 1/2n – минимальный шаг изменения частоты в популяции, см. также [3] для более строгого рассмотрения) и комбинируя (7), (8), мы получаем:
sdu /dP + (1/4n) d 2u /dP 2 = 0 . (10)
Решая это простое уравнение при естественных граничных условиях: u (1) = 1, u (0) = 0 , мы получим вероятность фиксации гена A1 в конечной популяции [1,3,6]:
u(P) = [1 - exp (- 4nsP)] [1 - exp (- 4ns)]-1 . (11)
Выражение (11) показывает, что если 4ns < < 1 , то имеет место нейтральная фиксация гена: u(P) » P , если 4ns > > 1, то отбирается предпочтительный ген A1 : u(P) » 1; размер популяции nc ~ (4s)-1 есть граничное значение, разделяющее области "нейтрального" и "селективного" отбора.
Итак, математические методы популяционной генетики описывают динамику частот генов в эволюционирующих популяциях. Детерминистические методы используются при описании динамики частот в среднем; стохастические методы учитывают флуктуации в популяциях конечной численности.
Похожие работы
... т.д. Зная эти закономерности, можно количественно исследовать генетическую структуру популяций и прогнозировать ее возможные изменения. Этому способствует солидная теоретическая база популяционной генетики – популяционно-генетические процессы математически формализованы и описаны уравнениями динамики. Для проверки различных гипотез о генетических процессах в популяциях разработаны статистические ...
нетика, микробиология, вирусология. Генетика человека — раздел генетики, изучающий закономерности наследования и изменчивости признаков у человека. Глава 2. Русские учёные в развитии генетики генетика наследственность ученый лобашев филипченко Филипченко Юрий Александрович У истоков отечественной генетики стояли выдающиеся ученые, которые пришли в новую науку из традиционных биологических ...
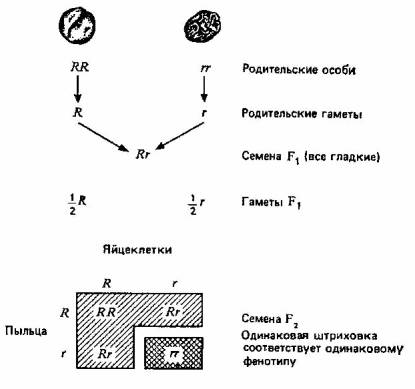
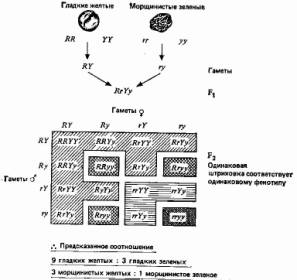

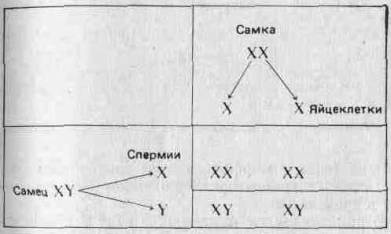
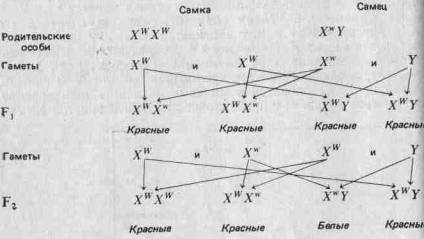
... , а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны. 3. Центральная догма и принцип Вейсмана принимаются. 4. Эволюция осуществляется путем изменения частот генов. 5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и ...
... другую. Получаются две двойные спирали – точные копии их предшественницы. Это свойство точно копировать себя имеет ключевое значение для жизни на Земле. 2. Генетика и медицина 2.1 Методы исследования В генетике основным методом исследования является генетический анализ, который проводится на всех уровнях организации живого (от молекулярного до популяционного). В зависимости от цели ...
0 комментариев