Вступление
Инсулин - полипептидный гормон, образованный 51 аминокислотами. Он секретируется в кровь b-клетками островков Лангерганса поджелудочной железы. Главная функция инсулина - регуляция метаболизма белков, жиров, углеводов. Это анаболический гормон. Его эффекты на мышцы, печень и адипозную ткань: стимуляция захвата клетками глюкозы, аминокислот, жирных кислот; усиление синтеза гликогена, белков, триглицеридов; стимуляция гликолиза; а также торможение глюконеогенеза и распада гликогена, белков и триглицеридов. Секреция инсулина минимальна при голодании, мышечной и нервной нагрузке, а также других формах стресса, когда возрастает потребность в использовании углеводов и жиров, и максимальна после приема пищи.
Секреция инсулина
Секреция инсулина контролируется изменениями концентраций циркулирующих в крови нутриентов (глюкозы, аминокислот, жирных кислот), гормонами желудочно-кишечного тракта, секретируемыми в нервно-гуморальную фазу сокоотделения (например, ГИП, гастрин, секретин) и различными нейромедиаторами (помимо классического ацетилхолина можно назвать такие пептидные медиаторы, как ВИП и холецистокинин). Перечисленные гормоны и медиаторы обуславливают так называемые энтероинсулярные стимулы секреции инсулина. Следует отметить, что их значение второстепенно; т. е. главными стимулами служат "пищевые" стимулы. По мере того, как концентрация, например, глюкозы в крови увеличивается [обычно достигая уровня 6-9 ммоль/л (норма: 5 ммоль/л)], стимулируется секреция инсулина, и этот эффект усиливается гормонами желудочно-кишечного тракта.
Показано, что эффекты нутриентов на секреторную активность b-клеток поджелудочной железы являются результатом их прямого взаимодействия с клеточными мембранами железистых клеток. Глюкоза и другие подвергающиеся метаболизму питательные вещества (включая некоторые аминокислоты и жирные кислоты) транспортируются в b-клетки островков Лангерганса, где в процессе их метаболизма образуется АТФ. Считается, что продукция АТФ обеспечивает стимул для начала секреции инсулина изменением мембранного потенциала, в конечном итоге обеспечивающим поток ионов Са2+ в цитоплазму.
В состоянии покоя мембранный потенциал (ПП) на внутренней поверхности мембраны равен -50-70мВ. Как известно, изменения ПП в большей степени контролируются изменением мембранной проницаемости для калия. В мембранах b-клеток существуют 2 типа калиевых каналов (АТФ-чувствительные и Са-чувствительные), оба из которых участвуют в секреции инсулина.
Образовавшийся АТФ вызывает закрытие АТФ-чувствительных калиевых каналов. Это предотвращает выход К+ из клетки, что является результатом накопления в ней положительных зарядов и, соответственно, деполяризации мембраны. По достижении порога (снижение потенциала на 15 мВ) открываются потенциал-чувствительные Са каналы, обеспечивая поток ионов Са2+ в клетки. Са-чувствительные калиевые каналы открываются по мере того, как Са2+ поступает в клетку, благодаря чему К+ выходит из нее, восстанавливая ПП.
Ионы Са2+ обеспечивают секрецию инсулина из секреторных гранул несколькими путями:
1) Положительно заряженные ионы Са2+ облегчают экзоцитоз (инсулин секретируется из клеток именно таким путем), уменьшая электростатическое отталкивание между отрицательно заряженными поверхностями плазматической мембраны и мембран секреторных гранул.
2) Са2+ облегчает передвижение гранул внутри клеток, т. к. влияет на функцию сократительных белков, содержащих актин и тубулин (микротрубочек и микрофиламентов).
3) Са2+ связывается с калмодулином; это активирует фермент аденилатциклазу, катализирующую превращение АТФ в цАМФ. Этот вторичный посредник также образуется в результате прямой активации АЦ гормонами желудочно-кишечного тракта. Циклический АМФ потенциирует секрецию инсулина путем увеличения чувствительности b-клеток к стимулирующему действию кальция. О клеточных процессах, лежащих в основе увеличения чувствительности b-клеток к Са2+, известно мало. Предполагается, что активируются ферменты (такие как протеинкиназы), влияющие на функционирование митротрубочек и микрофиламентов.
4) Чувствительность b-клеток к Са2+ увеличивается и другими вторичными мессенджерами (инозитолтрифосфатом и диацилглицеролом) предположительно таким же путем. Эти вторичные посредники образуются при взаимодействии нейромедиаторов энтероинсулярной оси (асh, холецистокинин) с фосфолипазой С, встроенной в плазматическую мембрану.
Еще раз следует подчеркнуть, что вышеперечисленные вторичные мессенджеры служат для увеличения секреции инсулина; тогда как главным стимулом служит увеличение концентрации глюкозы.
Действие инсулина на клетки
После секреции инсулина в межклеточное пространство он проходит через эндотелиальный барьер и попадает в кровь. Недавно стал известен механизм захвата глюкозы клетками. В этих клетках имеются небольшие везикулы, содержащие специфические белковые макромолекулы, которые называются транспортерами глюкозы. Инсулин стимулирует сплавление мембран везикул с плазматической мембраной и активирует транспортеры глюкозы, которые переносят глюкозу в клетку. Инсулин отвечает за синтез фермента гексокиназы, который фосфорилирует глюкозу, как только она проникает в клетку. Эффекты гормона на клеточном уровне достигаются путем активации или, наоборот, торможения активности ферментов или же изменением скорости синтеза ферментов на уровне транскрипции и трансляции.
Путь трансмембранного проведения гормонального сигнала - тирозинкиназный. Инициация действия инсулина обуславливается взаимодействием гормон-рецептор. Рецепторы инсулина относятся к семейству гетеротетрамерных рецепторов тирозинкиназы. Они образованы двумя парами a и b субъединиц, скрепленными дисульфидными мостиками. Две a субъединицы формируют один связывающий центр для лиганда. Образование Г-Р комплекса обуславливает аутофосфорилирование ферментов, "вмонтированных" в b субъединицы, по остаткам тирозина; в результате чего повышается ферментативная (тирозинкиназная) активность рецептора. Сигнал передается дальше посредством каскадных реакций:
1) Каскад ферментов (киназ и фосфатаз) приводит к усилению или торможению активности ферментов, обуславливающих эффекты инсулина, путем фосфорилирования или дефосфорилирования. Например, инсулин оказывает стимулирующий эффект на гликогенобразование, повышая активность гликогенсинтетазы (дефосфорилированием) и ингибируя гликогенфосфорилазу.
2) Эффекты инсулина, как уже упоминалось, могут также модулироваться изменением скорости синтеза ферментов на уровне транскрипции и трансляции. Этот путь включает в себя: фосфорилирование каскада МАР-киназ ® фосфорилирование с-myc (или c-fos) ® взаимодействие с-myc (или с-fos) с с-myc(с-fos)-зависимыми элементами ДНК ® изменение скорости синтеза ферментов.
Патологические процессы, связанные с нарушением молекулярных механизмов секреции инсулина и его действия на клетки
С глубокими нарушениями углеводного и жирового обмена у человека связана тяжелейшая эндокринная болезнь - сахарный диабет. В настоящее время считают, что в основе патогенеза диабета лежит сочетанное нарушение регулирующего действия инсулина и, возможно, ряда других гормонов на ткани; в результате чего в организме возникает абсолютная или относительная недостаточность инсулина, сочетающаяся с абсолютным или относительным избытком глюкагона или других "диабетогенных" гормонов.
Дисбаланс действия гормонов приводит соответственно к развитию устойчивой гипергликемии (концентрация сахара в крови выше 130 мг%), глюкозурии и полиурии. Последние два симптома и дали название заболеванию - сахарное мочеизнурение, или сахарный диабет. Наряду с нарушением утилизации и депонирования углеводов при диабете возникают различные расстройства жирового обмена, приводящие к гиперлипацидемии и, соответственно, повышенному образованию кетоновых тел (это обуславливает снижение рН-ацидоз).
Выделяют следующие формы диабета:
· При недостаточной секреции инсулина развивается инсулин-зависимая (инсулин-чувствительная) форма диабета.
· При резко сниженной чувствительности тканей-мишеней к инсулину развивается неинсулин-зависимая (инсулинрезистентная) форма.
Для лечения сахарного диабета применяют различные препараты инсулина (лишь при первой форме заболевания); малоуглеводную диету; сахароснижающие синтетические препараты-сульфанилмочевинные и бигуанидные (эти лекарства стимулируют секрецию инсулина и/или повышают чувствительность клеток-мишеней к гормону).
Таким образом, знание молекулярных механизмов секреции инсулина и действия его на клетки необходимо для выяснения, на каком уровне возник патологический процесс и какой путь лечения диабета будет эффективным.
Использованная литература:
à Peter R. Flatt and Clifford J. Bailey. “Molecular mechanisms of insulin secretion and insulin action”. Journal of Biological Education (1991) 25 (1)
à В. Б. Розен. Основы эндокринологии (1994)
Похожие работы
... взаимодействие с-myc (или с-fos) с с-myc(с-fos)-зависимыми элементами ДНК изменение скорости синтеза ферментов. Патологические процессы, связанные с нарушением молекулярных механизмов секреции инсулина и его действия на клетки С глубокими нарушениями углеводного и жирового обмена у человека связана тяжелейшая эндокринная болезнь сахарный диабет. В настоящее время считают, что в ...
... черепномозговых нервов; 2. . полинейропатия спинномозговых нервов: 3. дистальная полинейропатия (сенсо-моторная); 4. нейромиалгия (при декомпенсации сахарного диабета); 5. проксимальная амиотрофия; 6. автономная висцеральная нейропатия: 7. нейропатия органов кровообращения; 8. нейропатия органов ...
... , вызванные динамическими му-тациями.-----------------------T-----------T-------T-----T------T------T----------------------¬ Болезнь, номер по ¦ Ген, лока-¦Триплет¦Норма¦Прему-¦Мута- ¦Литература ¦ МакКьюсику (MIM) ¦ лизация ¦ ¦ ¦тация ¦ция ¦ ¦ -----------------------+-----------+-------+-----+------+------+----------------------+ Синдром ломкой X-хро- ¦FMR1, FRAXA¦(CGG)n ...
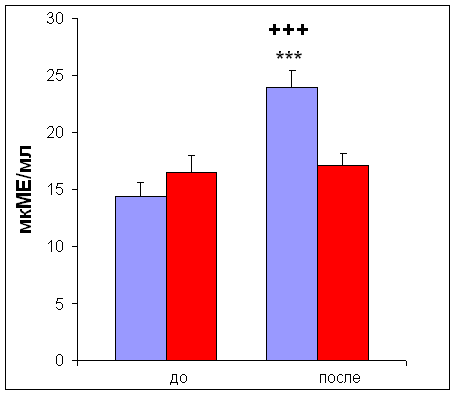
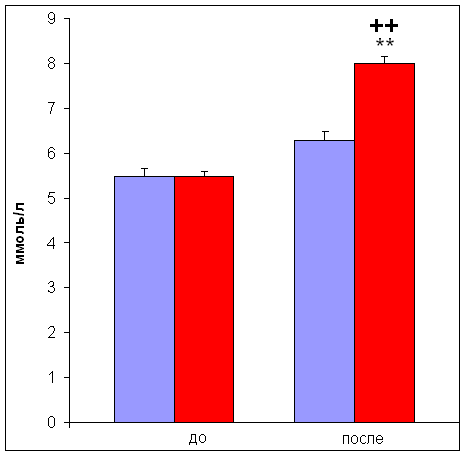
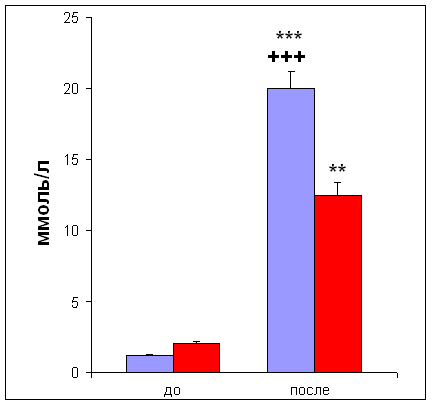
... м/с, на которой испытуемый бежал до состояния полного утомления [11]. 2.3. Определение уровня инсулина Уровень инсулина определяли иммуноферментным методом ELISA [23] в сыворотке крови. Для получения сыворотки кровь центрифугировали 30 мин при 4000 об/мин. 2.4. Определение уровня лактата и глюкозы Уровень глюкозы и лактата определяли с помощью ферментных электродов на автоматическом ...
0 комментариев