Навигация
Проявление старения на тканевом уровне
42969
знаков
1
таблица
1
изображение
5. Проявление старения на тканевом уровне.
[8]О существовании внутриклеточных факторов, ограничивающих продолжительность жизни клеток пределами, характерными для данного вида, свидетельствую и с исследования проведённые на культуре ткани.
Основатель метода Carrel в 1912г. ввёл в культуру ткани фибробласты сердечной мышцы зародыша цыплёнка путём непрерывных пассажей в свежую питательную среду фибробласты сохранялись в течении 34л. (Parken, 1961г.). Клетки всё время были молодыми и здоровыми и прожили тройной срок жизни цыплёнка, эквивалентной 200 годам жизни человека. Опыт был закончен в 1946г.
[9]С целью исследования причин старения организма было предпринято изучение старения клеток в культуре. Как впервые показали Свим и Паркер, фибробласты, полученные из различных тканей человека, пролиферируют только в течение ограниченного времени, после чего погибают. Эти авторы, однако, не выяснили, сохраняют ликлетки в культуре свои нормальные свойства, и не осмелились предположить, что их ограниченная пролиферативная активность связана со старением. Позже аналогичные работы были широко развернуты в лаборатории Хейфлика. Хейфлик и Мурхед установили, что легочные фибробласты эмбриона человека проходят в культуре около 50 удвоений популяции и затем погибают. Фибробласты, полученные из легочной ткани взрослого донора, подвергаются только ~20 удвоениям популяции. Хейфлик предположил, что имеется обратная зависимость между возрастом донора и пролиферативным потенциалом фибробластов. Позже другие исследователи также сообщили о наличии такой связи 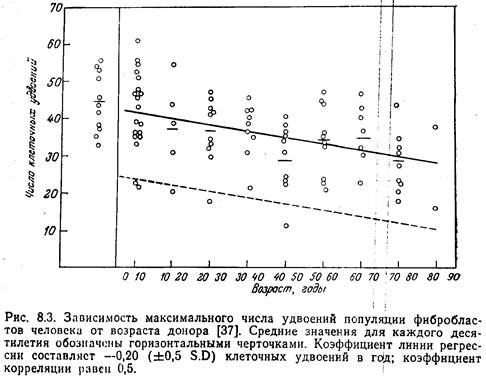
Более того, было показано, что при культивировании фибробластов кожи, полученных от людей в возрасте 10—90 лет, наблюдается снижение потенциала удвоения примерно на 0,20 относительные единицы в расчете на каждый год жизни донора. Коэффициент регрессии составляет —0,20. Сообщение Голдстейна и др. о том, что не хронологический возраст донора, а скорее его физиологическое состояние ответственно за скорость роста фибробластов, побудило многих авторов проделать скрупулезную работу с клетками разных типов, чтобы установить, действительно ли существует линейная связь между хронологическим возрастом и удвоением популяции. Изучение культур фибробластов, источником которых служили клетки тщательно отобранных здоровых доноров, не предрасположенных к диабету, дало отрицательный результат: обратная зависимость между возрастом донора и числом удвоений популяции не была обнаружена. В то же время эта зависимость была установлена для культур клеток, полученных от больных диабетом или от лиц, генетически предрасположенных к этому заболеванию. Таким образом, именно физиологическое состояние в большей степени, чем хронологический возраст, определяет пролиферативную активность клеток.
[10]Можно ожидать, что фибробласты больных прогерией (синдром Хатчинсона — Гилфорда) и с синдромом –Вернера, заболеваниями, для которых характерно ускоренное старение, должны иметь сниженную пролиферативную активность. Прогерия — редкое генетическое заболевание человека, при котором наблюдается замедление роста и атеросклероз всех крупных кровеносных сосудов, включая аорту и коронарную артерию, уже в 9-летнем возрасте, а также значительное отложение возрастного пигмента липофусцина в клетках многих органов. Картина преждевременного старения выражена очень ярко, и ребенок в возрасте 9 лет напоминает 70-летнего человека. Больные остаются бесплодными и умирают, не достигнув 20 лет. Синдром Вернера является сходным заболеванием, но его симптомы развиваются позже; больные тоже имеют сниженную продолжительность жизни, в среднем около 47 лет. Заболевание носит генетический характер, оно контролируется аутосомным геном. Основные симптомы; высокая частота злокачественных новообразований, ранняя потеря и поседение волос, малый, рост, юношеская катаракта, склонность к диабету, ранний атеросклероз, кальцификация кровеносных сосудов и остеопороз.
Фибробласты, полученные от 9-летних больных прогерией, проходят в культуре только 2—4 удвоения популяции, т. е. существенно меньше, чем клетки, полученные от здоровых детей того же возраста. Снижен синтез ДНК и эффективность клонирования. Антигенные маркеры HLA, имеющиеся на поверхности фибробластов здоровых людей, отсутствуют на фибробластах больных прогерией. Фибробласты больных синдромом Вернера также обладают- сниженным потенциалом удвоения. Эти данные показывают, что скорее физиологическое состояние, а не хронологический возраст определяет способность клеток к пролиферации. Более того, ускоренное старение организма отражается в ускоренном старении .клеток in vitro, о чем свидетельствует изменение скорости деления и метаболизма.
Известно, что если клетки заморозить при температуре ниже нуля после того, как они просуществовали в культуре некоторое время, например прошли 20 удвоений популяции, а затем вернуть их к жизни при 37 °С, то они возобновляют делёние и «доживают свой век», проходя еще около 30 удвоений популяции. Это указывает на (постоянство потенциала деления клеток данного типа, который не зависит от времени, необходимого для завершения полного цикла делений. Фибробласты человека штамма WI-38, замороженные в жидком азоте на 14 лет, после возобновления размножения завершили общий цикл деления, который составил 50±10 удвоений популяции.
Был измерен потенциал деления клеток различных типов. Мартин и др. показали, что фибробласты кожи эмбриона человека проходят 45 удвоений популяции, тогда как клетки скелетной мышцы и костного мозга—значительно меньше. Кроме того, выяснили, что клетки почки, сердца, тимуса, щитовидной железы и печени живут в культуре меньше, чем клетки легкого. Следовательно, пролиферативная активность клеток различных типов неодинакова. Зависит ли это от характера н степени их дифференцировки, пока неизвестно. Хотя клетки костного мозга и эпителия продолжают делиться в организме на протяжении всей жизни, их пролиферативная способность in vitro ограничена. В этой связи было бы важно узнать не только то, почему эти клетки имеют пониженный потенциал деления, но также и то, не делились ли они уже до помещения их в культуру. Такие исследования могут дать полезную информацию о том, почему клетки типа нейронов, скелетной и сердечной мышц прекращают деление на ранней стадии развития.
Были предприняты попытки установить взаимосвязь между продолжительностью жизни вида и потенциалом деления фибробластов, и хотя были изучены лишь немногие виды с разной .продолжительностью жизни, результаты указывают на наличие положительной корреляции между этими величинами.
Максимальная продолжительность жизни культивируемых нормальных фибробластов эмбриона человека и животных.
| Вид | Пределы удвоения популяции клеток | Максимальная продолжительность жизни, годы |
| Галапагосская черепаха | 90—125 | 175(?) |
| Человек | 40—60 | 110 |
| Норка | 30—34 | 10 |
| Курица | 15-35 | 30(?) |
| Мышь | 14—28 | 3,5 |
Фибробласты эмбриона галапагосской черепахи, наиболее долгоживущего вида среди изученных, обладают и самым высоким потенциалом деления, тогда как для мыши, чья продолжительность жизни не превышает трех лет, эта величина значительно меньше.
Для ответа на вопрос, вызваны ли старение и смерть клеток накоплением какого-либо цитоплазматического фактора, был использован метод гибридизации соматических клеток. Гибридизация старых и молодых клеток была нежелательна, так как она привела бы к образованию гибридов с двойным набором хромосом. Удалением ядра из легочных фибробластов человека, находившихся во II и III фазах роста, получали ци-топласты. Затем происходило их слияние с интактными фибробластами раннего и позднего пассажей следующим образом: молодые цитопласты—с молодыми клетками, старые цитопласты — с молодыми клетками, молодые цитопласты—со старыми клетками и старые цитопласты — со старыми клетками. Число удвоений популяции первых двух гибридных линий составило около 20, а двух последних — около 5. Отсюда следует, что цитоплазма, вероятно, не содержит никакого фактора, который влияет на потенциал деления, т. е. пролиферативная активность клеток определяется на уровне ядра или генома. Полученные результаты необходимо подтвердить в дальнейших исследованиях.
Таким образом, клетки стареют как in vivo, так и in vitro. Существует, однако, большая вариабельность продолжительности их жизни и числа делений. Клетки типа нейронов или скелетной и сердечной мышц прекращают деление in vivo на ранних стадиях развития и становятся постмитотическими. Было бы интересно узнать, сколько раз они подвергаются делению, прежде чем достигают постмитотической стадии. Спустя некоторое время эти постмитотические клетки стареют и погибают. Таким образом, эти клетки сначала прекращают деление, а затем подвергаются старению. Клетки костного мозга, эпителия и печени сохраняют пролиферативную активность в течение всей жизни, хотя она постепенно падает в старческом возрасте. Когда эти клетки культивируют in vitro, они прекращают делиться и через некоторое время стареют. Очевидно, эти клетки обладают способностью к размножению, которая во много раз превышает аналогичную способность нейронов и мышечных клеток. Важно изучить клетки эмбриона тотчас после того, как они прошли дифференцировку я превратились в клетки специфических типов, чтобы увидеть, имеют ли степень и тип дифференцировки какую-либо связь с пролиферативной активностью и природой постмитотического состояния. Такие исследования клеток различных типов, начиная с самых ранних стадий дифференцировки; и до прекращения деления, могут дать ключевую информацию о причинах прекращения деления.
Возникает вопрос, действительно ли те клетки, которые сохраняют способность делиться в течение всей жизни, не стареют? Действительно ли они обновляются всякий раз, когда делятся? Можно ли считать две дочерние клетки полностью идентичными не только в отношении содержания ДНК, но и родительских органелл? Известно, что во время созревания сперматиды наблюдается неадекватное деление
[11]Бернет предположил, что клетки одного типа могут быть более ответственны за старение, чём другие. По его мнению клетки тимуса и тимус-зависимые клетки, которые подвержены ослаблению пролиферации по типу фазы III, определяют темп старения. Бернет—сторонник той точки зрения, что изменения в этих клетках ответственны за аутоиммунные заболевания, которые чаще встречаются в пожилом возрасте. Эта концепция основана на представлении, что причиной старения являются соматические мутации, однако доказательств для нее пока нет.
Если старение—внутреннее свойство клеток, тогда оно должно быть присуще и одноклеточным организмам. Амебы проходят ограниченное число делений и живут ограниченное время, если их содержать на неполноценном рационе. Они продолжают расти и размножаться непрерывно только в том случае, если питание обильно, чего обычно не бывает в естественных условиях. Многие клопы парамеций и аскомицетов имеют ограниченную пролиферативную активность.
6.Литература1. Аршавский И. А. «Физиологические механизмы и закономерности индивидуального развития: основы негентропийной теории онтогенеза»- М:. : Наука, 1982г.
2. Лэмб М. «Биология старения»- М. : Мир, 1980г.
3. Стрелер Б. «Время, клетки и старение»- М. : Мир, 1964г.
4. «Физиологические механизмы старения» (С.А. Талин, А. С. Ступина, О. А. Мартыненко и др.)- Л. : Наука, 1982г.
5. Фрольксис В. В. «Природа старения. Биологические механизмы развития старения»- М. : Наука 1969г.
6. «Биология старения» (В.В. Фрольксис, И.А. Аршавский, Н.И. Арингин и др.)- Л. : Наука, 1982г.
7. Гериатрия (Учебное пособие). Под редакцией академика АМН СССР Д.Ф. Чеботарева- М, Медицина-1990г.
8. Аршавский И.А. «Очерки по возрастной физиологии»- М. : Медицина, 1967г.
9. Ванюшин Б.Ф., Бердышев Г.Д. «Молекулярно генетические механизмы старения»- М. : Медицина, 1977г.
10. Виленчик М.М. «Биологические основы старения и долголетия»- М.: Знание, 1976
[1] 6. с. 64-69
[2] 7. с. 12-16
[3] 4. с.190-193.
[4] 7. с.13-15
[5] 3. с.43-45.
[6] 8. с.120-124.
[7] 5. с.98-101.
[8]
1. [9] 9(с. 190-196)
2. 12(с. 238-243)
[10] 9. с.67-71.
[11] 10. с.137-140.
Похожие работы
... термостойкость резин на основе каучука Elarim 153 выше при использовании минеральных наполнителей. Сопротивление термическому старению таких резин снижается при добавлении ПТДХ и избытке вулканизующих веществ. Резины на основе фторкаучуков (ФК) Фторкаучуки - наиболее термо- и химически стойкие эластомеры. Максимальная температура длительной эксплуатации резин на основе каучуков типа СКФ-26 ...
... в области геронтологии. Это общая теория старения. Само собой разумеется, речь идет только об эскизах. Собственно это еще не общая теория старения, а только её очерк. Благодаря тому, что и как я сделал, моя скромная работа весьма отличается от известных до сих пор трудов по геронтологии. К сожалению, я не могу сразу предложить точные дефиниции, но, по крайней мере, имею возможность сделать ...


... в средней продолжительности жизни в европейских странах и развивающихся странах Азии Африки и Латинской Америки. Низкая продолжительность жизни во многих странах Африки и Азии - наследие длительного колониального режима со всеми его последствиями: низким уровнем материального обеспечения, нищетой, голодом или недоеданием, плохими жилищными и санитарно-гигиеническими условиями, тяжким физическим ...
... различных аутоантител, вырабатывающихся против собственных белков, значительно повышен. В период от 40 до 80 лет он может увеличиться в 6-8 раз. Все это ведет к саморазрушению и старению организма, его «иммунологическому разоружению». Критика этой гипотезы сводится к тому, что в этом случае речь идёт не о первичных изменениях. Поскольку сама иммунная система очень сложна, а ее регуляция не вполне ...
0 комментариев