Навигация
Ретикулярная формация ствола головного мозга
25034
знака
0
таблиц
2
изображения
Оглавление
Оглавление……………………………………………………………………………2
1. Анатомическое строение и волоконный состав…………………………3
2. Неспецифические нисходящие влияния………………………………….3
3. Восходящие влияния………………………………………………………5
4. Свойства ретикулярных нейронов………………………………………..8
5. Выводы……………………………………………………………………..10
Список литературы…………………………………………………………………...12
1. Анатомическое строение и волоконный состав
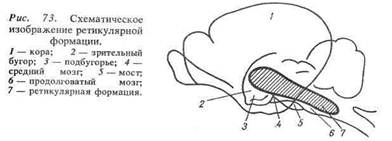
Ретикулярная формация - совокупность нейронов и соединяющих их нервных волокон, расположенных в стволе мозга и образующих сеть.
Ретикулярная формация простирается через весь ствол головного мозга от верхних шейных спинальных сегментов до промежуточного мозга. Анатомически она может быть разделена на ретикулярную формацию продолговатого мозга, варолиевого моста и среднего мозга. Вместе с тем, в функциональном отношении в ретикулярной формации разных отделов мозгового ствола есть много общего. Поэтому целесообразно рассматривать ее как единую структуру.
Ретикулярная формация представляет собой сложное скопление нервных клеток, характеризующихся обширно разветвленным дендритным деревом и длинными аксонами, часть из которых имеет нисходящее направление и образует ретикулоспинальные пути, а часть восходящее. В ретикулярную формацию поступает большое количество путей из других мозговых структур. С одной стороны, это коллатерали волокон, проходящих через ствол мозга сенсорных восходящих систем, эти коллатерали заканчиваются синапсами на дендритах и соме нейронов ретикулярной формации. С другой стороны, нисходящие пути, идущие из передних отделов мозга (в том числе, пирамидный путь), тоже дают большое количество коллатералей, которые входят в ретикулярную формацию и вступают в синаптические соединения с ее нейронами. Обилие волокон поступает к нейронам ретикулярной формации из мозжечка. Таким образом, по организации своих афферентных связей эта система приспособлена к объединению влияний из различных мозговых структур. Выходящие из нее пути могут оказывать в свою очередь влияния как на вышележащие, так и на нижележащие мозговые центры.
Нейронная организация ретикулярной формации до сих пор остается недостаточно изученной. В связи с чрезвычайно сложным переплетением в ней отростков различных клеток разобраться в характере межнейронных связей этой области очень трудно. Вначале широко распространенным было представление, что отдельные нейроны ретикулярной формации тесно связаны друг с другом и образуют нечто подобное нейропилю, в котором возбуждение распространяется диффузно, захватывая большое количество разных клеток. Однако результаты прямого изучения активности отдельных нейронов ретикулярной формации оказались не соответствующими таким представлениям. При микроэлектродном отведении такой активности выяснилось, что близкорасположенные клетки могут обладать совершенно различными функциональными характеристиками. Поэтому приходится думать, что организация межнейронных связей в ретикулярной формации достаточно дифференцирована и отдельные ее клетки соединены между собой довольно специфичными связями.
2. Неспецифические нисходящие влияния
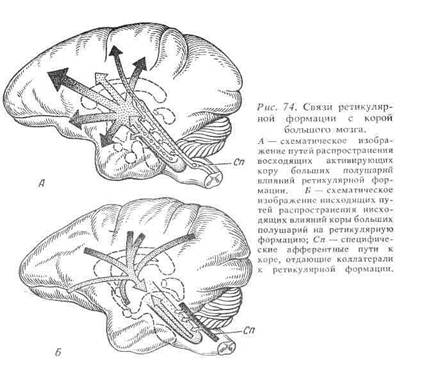
В 1946 г. американский нейрофизиолог X.Мегоун и его сотрудники обнаружили, что ретикулярная формация мозгового ствола имеет непосредственное отношение к регуляции не только вегетативной, но и соматической рефлекторной деятельности. Раздражая различные точки ретикулярной формации, можно чрезвычайно эффективно изменять течение спинальных двигательных рефлексов. В 1949 г. совместные работы X.Мегоуна и итальянского нейрофизиолога Дж.Моруцци показали, что раздражение ретикулярной формации эффективно влияет и на функции высших структур мозга, в частности коры больших полушарий, определяя переход ее в активное (бодрствующее) или неактивное (сонное) состояние. Эти работы сыграли исключительно важную роль в современной нейрофизиологии, поскольку они продемонстрировали, что ретикулярная формация занимает особое место среди других нервных центров, определяя в значительной мере общий уровень активности последних.
Влияния на двигательную деятельность спинного мозга возникают, в основном, при раздражении ретикулярной формации заднего мозга. Участки, которые создают эти эффекты, в настоящее время довольно хорошо определены, они совпадают с гигантоклеточным ядром ретикулярной формации продолговатого мозга и ретикулярным ядром моста. Эти ядра содержат большие клетки, аксоны которых образуют ретикулоспинальные пути.
Первые работы X.Мегоуна показали, что раздражение гигантоклеточного ядра вызывает ослабление в одинаковой мере всех спинальных двигательных рефлексов и сгибательных, и разгибательных. Поэтому он сделал вывод о том, что нисходящая система, берущая начало в вентрокаудальной части ретикулярной формации имеет неспецифическую тормозящую функцию. Несколько позже было обнаружено, что раздражение более дорсальных и оральных ее участков вызывало, наоборот, диффузное облегчающее действие на спинальную рефлекторную деятельность.
Микроэлектродные исследования эффектов, возникающих в нейронах спинного мозга при раздражении ретикулярной формации, действительно показали, что ретикулоспинальные влияния могут изменять передачу импульсации почти во всех рефлекторных дугах спинного мозга. Эти изменения оказываются очень глубокими и длительными, даже в том случае, когда ретикулярная формация раздражается всего несколькими стимулами, эффект в спинном мозге сохраняется сотни миллисекунд.
Одновременная активация большого количества ретикулоспинальных нейронов, которая имеет место в эксперименте с прямым раздражением ретикулярной формации и приводит к генерализованному изменению рефлекторной деятельности спинного мозга ситуация, конечно, искусственная. В естественных условиях такого глубокого сдвига этой деятельности, вероятно, не происходит; тем не менее, диффузное изменение рефлекторной возбудимости спинного мозга, несомненно, может иметь место при определенных состояниях мозга. Возможность диффузного ослабления рефлекторной возбудимости можно представить себе, например, при сне; оно приведет к снижению деятельности двигательной системы, характерному для сна. Важно учесть, что ретикулярное торможение захватывает также спинальные нейроны, участвующие в передаче афферентной импульсации в восходящем направлении, следовательно, оно должно ослаблять передачу сенсорной информации в высшие мозговые центры.
Синаптические механизмы диффузных влияний ретикулярной формации на нейроны спинного мозга изучены пока недостаточно. Как уже указывалось, эти влияния чрезвычайно длительны; кроме того, ретикулярное торможение устойчиво к действию стрихнина. Стрихнин является специфическим ядом, устраняющим постсинаптическое торможение мотонейронов, вызванное импульсацией из первичных афферентов и связанное, по всей вероятности, с выделением медиатора глицина. Нечувствительность диффузного ретикулярного торможения к стрихнину говорит, по-видимому, о том, что ретикулярные тормозящие эффекты создаются действием на спинальные клетки другого медиатора. Гистохимические исследования показали, что некоторое количество волокон в нисходящих путях, идущих из ретикулярной формации, является по своей природе адренэргическим. Однако пока неизвестно, имеют ли эти волокна отношение к диффузным ретикулоспинальным тормозящим эффектам.
Наряду с диффузными тормозящими влияниями, при раздражении определенных областей ретикулярной формации можно вызвать более специфические изменения деятельности спинальных элементов.
Если сравнить нисходящие влияния ретикулярной формации на нейронные структуры, регулирующие соматические и висцеральные функции, то можно обнаружить в них определенное сходство. Как сосудодвигательная, так и дыхательная функции ретикулярной формации строятся на сочетании деятельности двух реципрокно связанных между собой групп нейронов, оказывающих противоположное действие на спинальные структуры. Ретикулярные влияния на спинальные двигательные центры также состоят из противоположных, тормозящих и облегчающих компонентов. Поэтому создается впечатление, что реципрокный принцип организации нисходящих проекций представляет собой общее свойство ретикулярных структур; конечный эффект, соматический или вегетативный, определяется только тем, куда направлены аксоны соответствующих ретикулярных клеток. Такое сходство можно отметить и в других особенностях функционирования ретикулярных нейронов. Ретикулярные структуры, регулирующие вегетативные функции, отличаются высокой химической чувствительностью; влияния ретикулярной формации на двигательные центры также легко изменяются под влиянием таких химических факторов, как уровень CO2 в крови и содержание в ней физиологически активных веществ (адреналина). Механизм действия адреналина на ретикулярные нейроны довольно долго вызывал споры. Дело в том, что адреналин даже при его введении непосредственно в мозговую артерию может оказывать на ретикулярные нейроны непрямое действие (путем, например, сужения мозговых сосудов с последующей аноксией мозговой ткани). Однако исследование реакций ретикулярных нейронов в ответ на прямое приложение к ним адреналина через внеклеточный микроэлектрод показало, что некоторые из них действительно являются адреноцептивными.
Похожие работы

... чем нейроны других отделов мозга; тем не менее определенные их группы анатомически и физиологически тесно связаны с теми или иными специализированными отделами мозга. Микроскопическое строение коры головного мозга неодинаково в разных ее отделах. Эти морфологические различия коры позволили выделить отдельные корковые цитоархитектонические поля. Имеется несколько вариантов классификаций корковых ...
... месяцев с момента черепно-мозговой травмы. Сохраняются лишь астеническое состояние различной степени тяжести. Тяжесть и длительность психических нарушений в остром периоде травмы влияет на последующую реабилитацию больных. Травматическая астения при травме головного мозга, характеризуется снижением памяти, сужением круга интересов, повышенной возбудимостью и склонностью к аффективным вспышкам. ...
... мозга. Большая роль в осуществлении влияний мозжечка на функции организма принадлежит двусторонней связи мозжечка с моторной зоной коры большого мозга и ретикулярной формацией ствола мозга (рис. 76). Вегетативная нервная система Нервную систему организма животных и человека подразделяют на соматическую (анимальную) и вегетативную. Соматическая нервная система обеспечивает иннервацию ...
... мозга. Выявлены сложные реципроктные отношения между клетками ядер шва и ЦСВ, а также установлено наличие прямых проекций из ЦСВ в спинной мозг. Эти ядра традиционно рассматриваются как основное, конечное звено эндогенной болеутоляющей системы ЦСВ. В результате их возбуждения угнетается восходящий ноцицептивный поток, формируемый на сегментарном уровне. Наряду с нисходящей системой ядер шва, ...
0 комментариев