Навигация
Клинические проявления и иммунитет у птиц
63879
знаков
4
таблицы
0
изображений
2.2. Клинические проявления и иммунитет у птиц
Несмотря на высокую летальность вируса гриппа H5N1, большинство домашних кур в Гонконге не проявляли никаких клинических признаков болезни. В это же самое время в популяции кур циркулировал вирус гриппа H9N2. При изучении роли вируса H9N2 в защите кур от летальной вирусной инфекции H5N1 обнаружилось, что сыворотка от кур, инфицированных вирусом H9N2, не дает перекрестных реакций с вирусом H5N1 в реакции нейтрализации и реакции ингибирования гемагглютинации. Большинство кур, инфицированных вирусом гриппа H9N2 за 3-70 дней до контрольного заражения вирусом H5N1, выживали при контрольном заражении, однако инфицированные птицы выделяли вирус гриппа H5N1 в испражнениях. Адаптивный перенос Т лимфоцитов или Т-клеток CD81 от инбредных кур (B2/B2), инфицированных вирусом гриппа H9N2, к нативным инбредным курам (B2/B2) защищал их от летального вируса H5N1. In vitro анализ цитотоксичности показал, что Т-лимфоциты или Т-клетки CD81 от кур, инфицированных вирусом гриппа H9N2, распознают клетки-мишени, инфицированные как вирусом птичьего гриппа H5N1, так и H9N2 в зависимости от дозы. Это показывает, что перекрестный клеточный иммунитет, индуцированный вирусом гриппа H9N2, защитил домашних кур от летальной инфекции H5N1 в Гонконге в 1997 году, однако не предотвратил выделение вируса с фекалиями. Кроме того, это доказывает, что перекрестный клеточный иммунитет может изменить исход инфекции птичьего гриппа у домашней птицы и создать ситуацию сохранения вируса птичьего гриппа H5N1 (14).
Проводилось и сравнение различных вакцин. Три вакцины - вакцина на основе инактивированного целого вируса, вакцина на основе гемагглютинина вируса птичьего вируса, полученного из бакуловируса, и вакцина на основе рекомбинантного гемагглютинина вируса птичьего гриппа - были протестированы по их способности защищать цыплят против высокопатогенного вируса птичьего гриппа Н5. Вакцины и контрольные вирусы (или их белковые компоненты) были получены из полевых штаммов вируса птичьего гриппа различного происхождения и включали штаммы, полученные с 4 континентов, 6 видов хозяев и в течение 38-летнего периода. Вакцины защищали от проявления клинических симптомов и уменьшали количество вируса, выделяемого птицей, и титр вируса, выделяемого за введением гемагглютинина контрольного вируса птичьего гриппа Н5. Иммунизация этими вакцинами должна снизить распространение вируса птичьего гриппа через респираторный и пищеварительный тракт и уменьшить передачу инфекции от птицы к птице. Хотя наиболее значимое уменьшение выделения вируса через респираторный тракт достигалось, когда вакцина была наиболее схожа с контрольным вирусом, генетический дрейф вируса птичьего гриппа не должен влиять на основную защиту, как и в случае человеческого гриппа (15).
Инфекция среди домашней птицы может быть неочевидной или вызывать респираторные заболевания, уменьшение яйценоскости или быстрое фатальное системное заболевание, известное как высокопатогенный птичий грипп. Нейтрализующие антитела к белкам гемагглютинина и нейраминидазы обеспечивают первичную защиту против болезни. Различные вакцины вызывают выработку нейтрализующих антител, включая убитые цельновирионные вакцины и рекомбинантные вакцины на основе вируса вакцины. По-видимому, антигенный дрейф вируса в случае птичьего гриппа играет меньшую роль при неудачном вакцинировании по сравнению с гриппом человека. Цитотоксический Т-лимфоцитный ответ может уменьшать выделение вируса в окружающую среду в случае низкопатогенного птичьего гриппа, но обеспечивает спорную защиту против высокопатогенного птичьего гриппа. Вирус гриппа может напрямую влиять на иммунный ответ инфицированных птиц, однако роль МХ гена, интерферонов и других цитокинов в защите против птичьего гриппа остается неизвестной (16).
3. Характеристика птичьего гриппа у человека3.1. Случаи заболевания птичьим гриппом среди людей
1997 - в Гонконге, вирус птичьего гриппа (H5N1) инфицировал как кур, так и людей. Это был первый случай, когда обнаружилось, что вирус птичьего гриппа может напрямую передаваться от птиц человеку. В ходе этой вспышки 18 человек (9 детей и 9 взрослых) были госпитализированы и 6 из них (1 ребенок и 5 взрослых) погибли. Для ликвидации вспышки было уничтожено 1,5 миллиона кур с целью уничтожения источника вируса. Ученые определили, что вирус распространился напрямую от птиц к человеку, хотя не исключалась возможность переноса и от человека к человеку.
1999 - в Гонконге, вирус птичьего гриппа А H9N2 инфицировал двоих детей. Оба ребенка выздоровели, других случаев заболевания зарегистрировано не было. Исследования показали, что источником инфекции была домашняя птица, от которой вирус напрямую передался человеку. Однако возможность передачи вируса от человека к человеку полностью не исключалась. Кроме того сообщалось о нескольких случаях инфекции вирусом H9N2 среди людей в материковой части Китая в 1998-1999 годах.
2003 - два случая инфекции птичьего гриппа H5N1 произошли в Гонконгской семье, приехавшей из Китая. Один человек погиб, другой выздоровел. Где и как произошло заражение этой семьи, не установлено. Другой член этой семьи погиб от респираторного заболевания в Китае, но тестирование этого случая проведено не было. О других случаях заболевания не сообщалось.
2003 - Два случая инфекции вирусом птичьего гриппа H7N7 среди рабочих птицеферм и членов их семей были установлены в Нидерландах в ходе вспышки птичьего гриппа среди домашней птицы. Вирус был обнаружен у 86 человек, ухаживающих за зараженной птицей, и 3 человек - членов их семей (17). Симптомы чаще всего ограничивались инфекцией глаз с некоторыми симптомами респираторных заболеваний, однако 1 пациент погиб (ветеринар, посещавший пораженную ферму). Есть несколько предположительных случаев передачи вируса от человека человеку.
2003 - установлена инфекция H9N2 у ребенка в Гонконге. Ребенок был госпитализирован и выздоровел.
2004 - вспышка птичьего гриппа H5N1 среди людей:
|
В мае 1997г. 3-летний мальчик в Гонконге страдал от лихорадки, язвах в горле и кашля. Его болезнь продолжалась около 2 недель и он умер от пневмонии. Из трахеальной жидкости был выделен вирус гриппа А, однако он не мог быть типирован стандартными реагентами. Это заставило думать о новом штамме. В августе 3 лаборатории независимо идентифицировали этот вирус как штамм нового для людей гриппа А (H5N1). До заболевания мальчик имел контакт с инфицированными курами. Таким образом, это был первый подтвержденный документально случай инфекции вирусом А птичьего гриппа H5N1 у человека. До этого случая считалось, что вирус птичьего гриппа поражает только птиц. Затем инфекция этим же вирусом была подтверждена у 17 других пациентов в возрасте от 2 до 60 лет. К январю 1998 года 6 человек умерли от болезни. Прямых доказательств передачи вируса от человека к человеку нет: все инфицированные (даже живущие вместе в одной комнате) имели контакт с инфицированной птицей. Никаких вакцин для этого штамма нет, и в настоящее время предпринимаются попытки найти кандидатный вакцинный штамм для разработки и производства коммерческой вакцины (18).
Основные отличительные особенности вируса образца 2004 года кратко можно сформулировать следующим образом:
Вирус стал более вирулентным, что свидетельствует о мутации вируса.
Вирус преодолел межвидовой барьер от птиц к человеку, однако пока нет доказательств того, что вирус передается напрямую от человека к человеку (все заболевшие люди имели прямой контакт с зараженной птицей).
Вирус поражает и убивает в основном детей.
Источник заражения и пути распространения вируса не определены, что делает ситуацию с распространением вируса практически не контролируемой.
Меры по предотвращению распространения - полное уничтожение всего поголовья птицы.
Вспышка птичьего гриппа в Гонконге с особой отчетливостью продемонстрировала роль домашней птицы как источника инфекции для человека (19).
В мае 2001 года вирус гриппа А субтипа H5N1 был выделен из мяса уток, импортированного в Северную Корею из Китая. Несмотря на то, что этот изолят не был столь патогенен, как выделенный в 1997 году, сам факт выделения высокопатогенного вируса гриппа H5N1 из домашней птицы говорит о том, что циркуляция вируса в Китае продолжается и может представлять риск переноса вируса от птиц к человеку (20). Перманентная циркуляция вируса птичьего гриппа типов H5N1 и H9N2, преодолевших видовой барьер от птиц к человеку в 1997 и 1999, потенциально может вызвать пандемию среди людей (21). Однако, несмотря на то, что вирус птичьего гриппа обладает некоторыми признаками пандемичного вируса, он не обладает способностью быстро распространяться среди популяции людей (22), что является необходимым условием для возникновения пандемии.
Птичий вирус трудно остановить, поскольку вирус, по-видимому, мутировал со времени последней вспышки в Гонконге в 1997 и 2003 году. Мигрирующие птицы могут распространять его, что подтверждает тот факт, что в Гонконге был найден погибший обыкновенный сокол (сапсан), переносящий этот вирус (23).
В отличие от вируса 1997 и 2003 года вирус H5N1 образца 2004 года стал более вирулентным, о чем свидетельствует необычно большое количество погибшей домашней птицы. Это увеличивает риск заболевания людей. Необходимо обратить внимание и на увеличивающуюся опасность охлажденного и замороженного мяса домашней птицы, поскольку вирус H5N1 может сохраняться в течение многих лет при температуре ниже -700С. Однако он уничтожается при качественном приготовлении мяса (24).
Локализация вспышек птичьего гриппа зависит от точности идентификации способов распространения вируса. Необычно то, что он первично распространяется перелетными птицами. Из предыдущего опыта известно, что ответственными за распространение птичьего гриппа между фермами являются люди и оборудование. В 1997 году вспышку в Гон-Конге удалось локализовать благодаря тому, что было уничтожено все поголовье домашней птицы в стране. В настоящее время вирус распространился среди домашней птицы по всей Азии, поэтому локализовать вспышку гораздо сложнее (25).
По сравнению с предыдущими вспышками, эпидемия птичьего гриппа 2004 года может охватить гораздо большее число ферм. Одновременно возможна трансмиссия вируса через Азию, так как не контролируются факторы, вызывающие распространение вируса. ВОЗ отмечает, что почти одновременные вспышки птичьего гриппа в Японии, Северной Корее, Вьетнаме, а теперь и в Таиланде и Камбодже исторически беспрецедентны и существует опасение, что этот новый, вирулентный штамм вируса птичьего гриппа может поразить весь мир.
Скорость эволюции вируса птичьего гриппа в природных хозяевах (водоплавающие птицы, ржанки и чайки) и аберрантных хозяевах (куры, индейки, поросята, лошади и люди) различается. Скорость эволюции, определенная для всех трех вспышек, была сходной со скоростью, наблюдаемой у млекопитающих, что служит веским доказательством адаптации вируса птичьего гриппа к новым видам хозяев (26). Пока что, по-видимому, птичий грипп не передается от человека к человеку, однако из-за эпидемии среди домашней птицы такая передача становится все более вероятной. Необходима всего лишь правильная рекомбинация между штаммом H5N1 и сосуществующим штаммом гриппа человека. Это может произойти, если кто-либо из людей или других животных заболеет человеческим и птичьим гриппом одновременно, что позволит вирусам обменяться генами и образовать новый штамм, который сможет легко передаваться от человека к человеку. До сих пор нет доказательств, что это произошло, поскольку во всех известных случаях болезни инфицирование происходило при прямом контакте с курами. Данная ситуация опасна тем, что если возникнет пандемия, она будет более трагична по последствиям, чем пандемия 1968 года.
Птичий грипп поражает в основном детей - по сообщениями агентства Рейтер на 26.01.2004г. из 7 жертв птичьего гриппа 6 - дети. Почему это происходит, неизвестно (27).
3.3. Клинические проявления, патогенезСимптомы птичьего гриппа у человека варьируют от типичных гриппо-подобных симптомов (лихорадка, кашель, боль в горле и боль в мышцах) до инфекции глаз, пневмонии, острого респираторного заболевания, вирусной пневмонии и других тяжелых, угрожающих жизни симптомов.
Патогенез вируса птичьего гриппа изучался на мышах, поскольку это одна из наиболее широко применяемых и изученных моделей для исследования патогенеза вирусов на млекопитающих, однако в качестве альтернативы предлагается изучать вирус птичьего гриппа на хорьках, для которых он также является патогенным (28).
Исследование патогенности птичьих и человеческих изолятов вируса H5N1 из Гонконга на 6-8 недельных мышах BALB/c показало, что как птичьи, так и человеческие изоляты вызывали у мышей заболевание, характеризующееся гипотермией, клиническими симптомами, быстрой потерей веса и 75-100% смертностью на 6-8 день после инфицирования. Три изолята не из Гонконга не давали никаких клинических проявлений. Один изолят A/tk/England/91 (H5N1) вызывал заболевание средней тяжести и все животные, кроме одного, выздоровели. Инфекция приводила к поражению (от легкого до тяжелого) как верхних, так и нижних отделов респираторного тракта. Наиболее часто вирус вызывал некрозы в респираторном эпителии полости носа, трахеи, бронхах и бронхиолах с сопутствующим воспалением. Наиболее серьезные и обширные поражения наблюдались в легких мышей, инфицированных гонконгским вирусом птичьего гриппа, в то время как у мышей, инфицированных вирусами A/ck/Scotland/59 (H5N1) и A/ck/Queretaro/95 (H5N2), поражения были слабыми или не наблюдались вовсе. Вирусы A/ck/Italy/97 (H5N2) и A/tk/England/91 (H5N1) проявили промежуточную патогенность, давая поражения респираторного тракта от слабых до средних. Кроме того, инфекция, вызванная различными изолятами вируса, могла быть в дальнейшем определена по иммунному ответу мышей. Изоляты не гонконгского происхождения после инфицирования вызывали продуцирование повышенных уровней активного трансформирующего фактора роста b, в то время как гонконгские изоляты такого не вызывали (29).
При заражении мышей человеческим изолятом вируса гриппа А H5N1, выделяются две группы, отличающиеся по вирулентности. Используя современные методы генетики, было показано, что мутация в позиции 627 в белке РВ2 влияет на исход инфекции у мышей. Более того, высокая расщепляемость гемагглютина является необходимым условием летальности инфекции (30).
Более ранние исследования также указывали на наличие двух групп вирусов: группа 1, для которой MLD50 находилась между 0.3 и 11 PFU, и группа 2, для которой MLD50 была более 103 PFU. Спустя день после интраназальной инокуляции мышам 100 PFU вируса группы 1, титр вируса в легких составлял 107 PFU/g или на 3 log больше, чем для вирусов второй группы. Оба типа вирусов реплицировались до высоких титров (>106 PFU/g) в легких на 3 день и оставались на этом уровне в течение 6 дней. Более важно то, что только вирусы первой группы вызывали системную инфекцию, реплицировались в нереспираторных органах, включая головной мозг. Иммуногистохимический анализ показал, что репликация вирусов первой группы происходила в нейронах головного мозга, глиальных клетках и кардиальных миофибрах.
Механизм вирулентности, ответственный за летальность вирусов гриппа у птиц, действует также и у хозяев-млекопитающих. То, что некоторые вирусы H5N1 не продуцировали системной инфекции на моделях, свидетельствует о том, что множественные факторы, которые еще должны быть установлены, вносят свой вклад в тяжесть H5N1 инфекции у млекопитающих. Кроме того, способность этих вирусов продуцировать системную инфекцию на мышах и отчетливые отличия в патогенности среди изолятов, показывает, что эта система является полезной моделью для изучения патогенеза вируса птичьего гриппа на млекопитающих (31).
Кроме того, показано, что одним из факторов, оказывающих влияние на патогенез вируса H5N1, является деструктивное влияние на иммунную систему, которое отличается у летальных и нелетальных изолятов вируса H5N1 (32).
Биохимическим аспектам, которые влияют на вирулентность, адаптацию вируса к новому хозяину, иммунный ответ и патогенез, посвящен целый ряд работ (33, 34, 35, 36, 37, 38, 39, 40).
3.4. ВакциныСразу же после вспышек 1997-1999 годов начались поиски вакцины против вируса птичьего гриппа. Поскольку неадаптированный H5N1 вирус является патогенным для мышей, именно эти животные были использованы как модель иммунной системы млекопитающих для исследования летальной инфекции птичьего гриппа.
Производство вакцины против вируса гриппа H5N1 в системе куриных эмбрионов невозможно из-за гибели куриных эмбрионов при заражении этим вирусом и высоком уровнем биобезопасности, необходимым для работы с этим вирусом и производством вакцины на основе этого вируса. Для разработки вакцины на основе цельного вируса использовались авирулентный вирус H5N4, выделенный от мигрирующих уток, вирус H5N1 и авирулентный рекомбинантный вирус H5N1. Все вакцины были инактивированы формалином. Интраперитональная иммунизация мышей каждой вакциной вызывала выработку гемагглютинин-ингибирующих и вирус-нейтрализующих антител, в то время как интраназальная вакцинация без адьюванта индуцировала как мукозальный, так и системынй антительный ответ, который защищал мышей от контрольного заражения летальным вирусом H5N1 (41).
Интрамускулярное введение вакцины, приготовленной на основе непатогенного штамма A/Duck/Singapore-Q/F119-3/97 (H5N3), антигенно связанного с человеческим вирусом H5N1, в сочетании с квасцами или без них, приводило к полной защите от летального контрольного заражения вирусом H5N1. Защита от инфекции наблюдалась у 70% животных, которым вакцина вводилась сама по себе и у 100% животных, которым вакцина вводилась в сочетании с квасцами. Протективный эффект вакцинации коррелировал с уровнем вирус-специфических сывороточных антител. Эти результаты говорят о том, что в случае пандемии возможно использование антигенно связанных, но не патогенных вирусов гриппа в качестве кандидатов в вакцину (42, 43).
Исследования ДНК вакцин показали, что ДНК вакцина, кодирующая гемагглютинин из A/Ty/Ir/1/83 (H5N8), который отличается от A/HK/156/97 (H5N1) в пределах 12% в НА1, предотвращает гибель мышей, но не заболевание при инфицировании H5N1. Следовательно, ДНК вакцина, сделанная на основе гетерологичного штамма Н5 не защищает мышей от инфицирования вирусом птичьего гриппа H5N1, но полезна при защите мышей от гибели (44).
Противогриппозные вакцины, индуцирующие значительный перекрестный гетеросубтипный иммунитет, могут преодолеть ограничения эффективности вакцин, вызванные антигенной вариабельностью вируса гриппа А. Мыши, получившие трехкратную интраназальную иммунизацию вакциной H3N2 в сочетании LT(R192G), были полностью защищены при летальном контрольном инфицировании высокопатогенным человеческим вирусом H5N1, причем вирусные титры в носовой полости и легких были по крайней мере в 2500 раз ниже, чем у контрольных мышей, получавших только LT(R192G). Напротив, мыши, которые получили трехкратную вакцинацию вакциной H3N2 подкожно в присутствии или отсутствии LT(R192G) или неполного адьюванта Фрейнда, не были защищены при летальном контрольном инфицировании и никакого заметного снижения титров вируса в тканях не наблюдалось на 5 день после контрольного инфицирования вирусом H5N1. Вакцинация без LT(R192G) приводила лишь к частичной защите против гетеросубтипного контрольного заражения. Результаты исследования гетеросубтипного иммунитета подтвердили полезность мукозальной вакцинации, которая стимулирует перекрестную защиту против множества вирусных субтипов, включая вирусы, представляющие потенциальную пандемическую опасность (45).
3.5. Разработка средств обнаружения и диагностикиВо время вспышки 1997 года анализ ингибирования гемагглютинации, стандартный для серологического определения инфекции гриппа у человека, показал низкую чувствительность при определении антител к вирусу птичьего гриппа. В связи с этим, для определения антител к вирусу птичьего гриппа у человека был предложен более чувствительный метод микронейтрализации и Н5 специфический непрямой ELISA (иммуноферментный анализ). Чувствительность и специфичность этих методов была сравнима и, кроме того, значительно увеличивалась при сочетании с Вестерн-блотом. Максимальная чувствительность (80%) и специфичность (96%) при определении анти Н5 антител у взрослых в возрасте от 18 до 59 лет достигалась при применении микронейтрализации в сочетании с Вестерн блотом, а максимальная чувствительность (100%) и специфичность (100%) при определении анти Н5 антител в сыворотке детей моложе 15 лет достигалась при применении ELISA в сочетании с Вестерн блотом. Этот алгоритм может использоваться при проведении сероэпидемиологических исследований вспышек птичьего гриппа H5N1 (46).
Было также показано, что высокопатогенные нейротропные варианты вируса птичьего гриппа H5N1 могут быть быстро выделены на мышах (47).
Кроме того, еще в 1995 году для быстрого определения последовательности сайта расщепления гемагглютинина, маркера потенциала вирулентности вирусов птичьего гриппа, была использована RT-PCR (полимеразная цепная реакция). Эта методика в сочетании с сиквенсом сайта расщепления гемагглютинина может служить в качестве быстрого и чувствительного метода оценки потенциальной вирулентности вирусов птичьего гриппа. Раннее обнаружение связанных с вирулентностью последовательностей на сайте расщепления гемагглютинина в полевых изолятах вируса поможет лучше контролировать грипп среди огромной популяции домашней птицы (48).
В дальнейшем был разработан простой молекулярный метод быстрого генотипирования для мониторинга внутренних генов циркулирующего вируса гриппа А. Стратегия субтипирования вируса была протестирована вслепую на 10 контрольных вирусах каждого субтипа H1N1, H3N2 и H5N1 (всего на 30) и обнаружила высокую эффективность. Стандартизованный метод генотипирования использовался для идентификации источника внутренних генов 51 вируса гриппа А, выделенного от людей в Гонконге в ходе вспышек 1997-1998 годов и сразу после них. Эта же методика использовалась для характеристики внутренних генов двух изолятов вируса птичьего гриппа H9N2, полученных в Гонконге в 1999г. (49)
Позднее был разработан real-time reverse transcriptase PCR (RRT-PCR) анализ для быстрого определения вируса гриппа А и субтипов Н5 и Н7 вируса гриппа А. В этом анализе используется одностадийный способ определения и флуоресцентные зонды. Предел определения - около 1000 копий мишени-РНК. С помощью этого метода можно определить 0,1 50%-ную инфекционную дозу для куриных эмбрионов. Для анализа субтипов вируса гриппа А предел определения - 103-104 копии мишени-РНК. Чувствительность и специфичность данного метода напрямую сравнивалась со стандартными методиками для определения вируса гриппа: выделение гриппа на куриных эмбрионах и субтипирование гемагглютинина в реакции ингибирования гемагглютинации. Сравнение проводилось на 1550 трахеальных и клоачных мазках от различных видов птиц и мазков, взятых из окружающей среды на рынках живой птицы в Нью-Йорке и Нью-Джерси. Результаты RRT-PCR коррелировали с результатами выделения гриппа на куриных эмбрионах в 89% образцов. Остальные образцы были положительными при определении только одним из методов. В целом чувствительность и специфичность Н7- и Н5-специфичных анализов была сходна с методом выделения вируса на куриных эмбрионах и реакции ингибирования гемагглютинации (50).
3.6. Лечение заболеванияИсследования, проводимые до сих пор, подтверждают, что назначение лекарств, разработанных для штаммов человеческого гриппа, будут эффективны и в случае инфекции птичьего гриппа у человека, однако не исключена возможность, что штаммы гриппа могут стать резистентными к таким лекарствам и эти лекарства станут неэффективными.
Было обнаружено, что выделенный вирус чувствителен к амантадину и римантадину, ингибирующим репродукцию вируса гриппа А и применяемым в терапии человеческого гриппа (51). Кроме того, был исследован ряд других препаратов. Ингибитор нейраминидаз занзивир ингибировал репликацию вирусов на клетках почек хомяков в анализе вирусного урожая (50% эффективная концентрация, 8,5-14,0 mM) и ингибировал активность вирусной нейраминидазы (50% ингибирующая концентрация 5-10 nM). Интраназальное введение занзивира дважды в день (50 и 100 mg/kg веса тела) полностью защищало мышей от смерти. В дозе 10 mg/kg веса занзивир полностью защищал мышей от инфицирования вирусом H9N2 и увеличивал продолжительность жизни и количество выживших мышей, инфицированных вирусами H6N1 и H5N1. Во всех исследованных дозах занзивир значительно уменьшал титры вируса в легких и полностью блокировал распространение вируса в мозг. Таким образом, занзивир является эффективным при лечении птичьего гриппа, который может быть перенесен на млекопитающих (52).
Орально вводимый ингибитор нейраминидазы RWJ-270201 был протестирован в параллели с занамивиром (zanamivir) и озелтамивиром (oseltamivir) на панели вирусов птичьего гриппа на ингибирование нейраминидазной активности и репликации в тканевых культурах. Затем эти агенты были протестированы на защиту мышей против летальных инфекций H5N1 и H9N2. In vitro, RWJ-270201 был наиболее эффективным против всех девяти субтипов нейраминидазы. RWJ-270201 (концентрация 50% ингибирования от 0,9 до 4,3 nM) превосходил занамивир и озельтамивир карбоксилат по ингибированию нейраминидазы. RWJ-270201 ингибировал репликацию вируса птичьего гриппа как евразийской, так и американской линии на клетках MDCK (концентрация 50% эффективности от 0,5 до 11,8 mM). Мыши, которым ежедневно давали RWJ-270201 из расчета 10 mg на кг веса были полностью защищены против контрольного заражения летальной дозой вирусов A/Hong Kong/156/97 (H5N1) и A/quail/Hong Kong/G1/97 (H9N2). Как RWJ-270201, так и озельтамивир значительно уменьшали титры вирусов в легких мышей при дневных дозах от 1,0 до 10 mg/kg и защищали распространение вируса в мозг. Когда лечение начиналось через 48 часов после экспозиции вирусом H5N1, 10 mg RWJ-270201/kg веса ежедневно защищали 50% мышей от гибели. Эти результаты подтвердили, что RWJ-270201 эффективен против вируса птичьего гриппа по крайней мере также, как и занамивир или озельтамивир и потенциально может использоваться в клинической практике для лечения птичьего гриппа при переносе его от птиц к человеку (53).
4. Потенциальная опасность пандемии гриппаВсе вирусы гриппа обладают потенциальной способностью изменяться. Существует возможность того, что вирус птичьего гриппа может измениться таким образом, что сможет инфицировать людей и легко распространяться от человека к человеку. Поскольку эти вирусы обычно человека не инфицируют, в человеческой популяции существует очень низкая иммунная защита против таких вирусов или эта защита отсутствует вовсе. В случае, если вирус птичьего гриппа станет способным инфицировать людей и приобретет способность легко распространяться от человека к человеку, может начаться пандемия гриппа. Этот факт подтверждают американские и британские ученые в своем сообщении 05 февраля 2004 года: результаты их исследований свидетельствуют о том, что испанский грипп был настолько смертельным из-за того, что он эволюционировал из птичьего гриппа и содержал уникальный белок, к которому у человека не было иммунитета (54). Об этом же свидетельствуют и данные по степени дивергенции антигенных сайтов гемагглютинина в ходе антигенного дрейфа вируса между 1918 и 1934 годами, подтверждающие гипотезу о том, что вирус человеческого гриппа, вызвавший пандемию 1918 года произошел от вируса птичьего гриппа подтипа Н1, который преодолел видовой барьер от птиц к человеку и адаптировался к человеку, предположительно путем мутации и/или реассортации незадолго до 1918 года (55).
Обычно вирусы гриппа А имеют четко определенный круг хозяев, однако ограничение круга хозяев полигенно по своей природе и не имеет абсолютного характера (11, 56). Иногда происходит межвидовая передача вируса как в природных условиях, так и при адаптации к новому хозяину в лабораторных условиях (11).
Для вирусов гриппа свойственна постоянная антигенная изменчивость. Два вида изменчивости - дрейф и сдвиг - меняют оба поверхностных антигена вируса гриппа А. При антигенном дрейфе происходят небольшие изменения в структуре гемагглютинина и нейраминидазы, в то время как при антигенном сдвиге изменения этих белковых молекул, вызванные реассортацией геномных сегментов, весьма значительны.
Ряд генетических и серологических данных свидетельствует о том, что пандемии гриппа человека могут быть результатом реассортации генов между вирусами человека и птиц. Это означает, что когда 2 вируса инфицируют одни и те же клетки, вирусное потомство может унаследовать наборы геномных РНК-сегментов, представляющие собой рекомбинации РНК-сегментов обоих родительских вирусов. Теоретически возможное число таких комбинаций, которые могут сформировать полный РНК-геном при конкурентной инфекции, составляет 2256. Однако лишь немногие вирусы-реассортанты обладают правильным сочетанием генов, необходимым для эффективной репродукции в природных условиях (11).
Генетические и биологические исследования подтверждают, что свиньи могут стать своеобразным "смешивающим сосудом" для образования нового реассортанта вируса гриппа, аналогичного пандемичным вирусам 1957 и 1968 годов (57).
В настоящее время возникновение пандемичного вируса гриппа возможно путем переноса генов из резервуара водоплавающих птиц к человеку через реассортацию в свиньях, гипотетическом "смешивающем сосуде". Понимание вспышки гриппа H5N1 в 1997 году в Гонконге и выделение вируса птичьего гриппа H9N2 от человека увеличивают альтернативные возможности для возникновения нового пандемичного вируса. Вирусы H9N2, обнаруженные в земноводной домашней птице в Южном Китае переместились обратно к водоплавающим домашним уткам, в которых эти вирусы генерируют множественные реассортанты. Эти новые вирусы H9N2 являются двойными или даже тройными реассортантами, которые обладают потенциальной способностью напрямую инфицировать людей. Некоторые из них содержат сегменты генов, которые полностью родственны таковым у A/Hong Kong/156/97 (H5N1/97, H5N1) или A/Quail/Hong Kong/G1/97 (G1-like, H9N2). Более важно то, что некоторые из этих внутренних генов полностью связаны с аналогичными генами нового вируса H5N1, выделенного в ходе вспышки в Гонконге в 2001 году. Обнаружена двухходовая трансмиссия вируса гриппа между земными и водными птицами, которая облегчает генерацию новых реассортантов вируса гриппа H9N2. Такие реассортанты могут играть прямую роль в появлении следующего пандемичного вируса (58). Вирусы H5N1 и H9N2 имеют сходные характеристики, что увеличивает вероятность появления нового патогена для человека (59). В материковой части Китая циркулируют гены, кодирующие H5N1, что сохраняет возможность вирусной реассортации (60). Вирус H5N1, циркулирующий на рынках живой птицы, охватывает две разных филогенетических линии во всех генах, которые очень быстро эволюционируют (61).
В соответствии с руководством ВОЗ, Министерство здравоохранения, социального обеспечения и спорта Нидерландов разработало национальный план по минимизации последствий пандемии гриппа. В рамках плана готовности к пандемии, была дана оценка значимости проблемы, исходя из количества госпитализированных и летальных случаев в ходе пандемии гриппа. Используя анализ сценария, были исследован и потенциальный эффект возможного вмешательства. Описываются и сравниваются сценарии развития для понимания потенциального воздействия пандемии (заболеваемости, госпитализации и гибели), различных видов вмешательства и критические параметры модели. Анализ сценариев является полезным инструментом для принятия политический решений, касающихся разработке и планирования контроля и управления эпидемией на национальном, региональном и локальном уровнях (62).
5. Пандемии гриппа в человеческой популяцииПандемия гриппа - это глобальная вспышка гриппа и происходит она, когда новый вирус гриппа появляется, распространяется и вызывает болезнь по всему миру. Последние пандемии вируса гриппа приводили к высоким уровням заболеваемости, смертности, социальной нестабильности и экономическим потерям.
В ХХ веке наблюдались три пандемии и 1 глобальная эпидемия, близкая к пандемии (1977). Возбудители пандемий распространялись по всему миру в течение примерно одного года после того, как были обнаружены.
Это:
1918-1919 - Испанский грипп, испанка [A (H1N1)]. Вызвал наибольшее количество смертей, более 500 000 людей погибли в США и от 20 до 50 миллионов людей погибли во всем мире. Множество людей погибли в течение первых нескольких дней после заболевания и множество - в результате осложнений после гриппа. Около половины погибших были молодые здоровые взрослые люди.
1957-1958 - Азиатский грипп [A (H2N2)]. Вызвал около 70 000 смертей в США. Впервые зарегистрирован в Китае в конце февраля 1957 года, азиатский грипп достиг США в июне 1957.
1968-1969 - Гонконгский грипп [A (H3N2)]. Вызвал около 34 000 смертей в США. Был впервые зарегистрирован в Гонконге в начале 1968 года и достиг США в конце этого же года. Вирус гриппа А (H3N2) циркулирует до сих пор.
Вирус гриппа впервые был выделен в 1933 г. Интересно, что каждый новый вирус (азиатский, гонконгский) сначала появлялся в Китае, и полагают, что вирусы, вызывавшие эпидемии, происходившие до 1933 г., также происходили из Китая (63).
Эти пандемические вирусы имели несколько общих особенностей. Первые вспышки пандемий, вызванные этими вирусами, произошли в Юго-Восточной Азии. Появление вирусов Н2N2 и Н3N2 сопровождалось исчезновением из человеческой популяции вирусов, циркулировавших до них (соответственно вирусов подтипов Н1N1 и Н2N2). Почему вирусы, ранее циркулировавшие в человеческой популяции, исчезали с появлением новых вирусов, остается неясным (11).
По антигенной специфичности пандемичные вирусы, ответственные за азиатский и гонконгский грипп, отличались от вирусов гриппа, циркулировавших у людей перед их появлением. Агент эпидемии "русского гриппа" в 1977 г. (подтип Н1N1) был в основном идентичен вирусам, циркулировавшим среди людей в 1950 г. В высшей степени сомнительно, что этот вирус сохранялся в природе более 20 лет без каких-либо изменений. Поэтому логично сделать заключение о том, что вирус был сохранен в замороженном виде до момента внедрения каким-то образом в человеческую популяцию (11).
Обычно, однажды появившись и распространившись, вирус гриппа обоснуется среди людей и циркулирует в течение многих лет. Центр США по контролю заболеваний и ВОЗ осуществляют обширные программы мониторинга случаев заболевания гриппом по всему миру, включая появление потенциально пандемичных штаммов вируса гриппа (64).
Список литературы
Swayne DE; Perdue ML; Garcia M; Rivera-Cruz E; Brugh M, Pathogenicity and diagnosis of H5N2 Mexican avian influenza viruses in chickens, Avian Dis 1997, 41(2):335-46.
Swayne DE; Beck JR; Mickle TR, Efficacy of recombinant fowl poxvirus vaccine in protecting chickens against a highly pathogenic Mexican-origin H5N2 avian influenza virus, Avian Dis 1997, 41(4):910-22.
Stallknecht ED. Ecology and epidemiology of avian influenza viruses in wild bird populations: waterfowl, shorebirds, pelicans, cormorants, etc., Proc. 4th International Symp. on Avian Influenza, May 29-31, 1997, Athens, USA, pp. 61-67.
Webster RG, Yakhno MA, Hinshaw VS, Bean WJ, Murti KG. Intestinal influenza: replication and characterization of influenza viruses in ducks. Virology 1978; 84:268-78.
Geraci JR, St. Aubin DJ, Barker IK, Webster RG, Hinshaw VS, Bean WJ, et al. Mass mortality of harbor seals: pneumonia associated with influenza A virus. Science 1982; 215: 1129-31.
Hinshaw VS, Bean WJ, Geraci JR, Fiorelli P, Early G, Webster RG. Characterization of two influenza A viruses from a pilot whale. J Virol 1986; 58:655-6.
Scholtissek C, Burger H, Bachmann PA, Hannoun C. Genetic relatedness of hemagglutinins of the H1 subtype of influenza A viruses isolated from swine and birds. Virology 1983; 129:521-3.
Englund L, Klingeborn B, Mejerland T, Avian influenza A virus causing an outbreak of contagious interstitial pneumonia in mink, Acta Vet Scand 1986; 27(4):497-504.
Guo Y, Wang , Kawaoka Y, Gorman O, Ito T, Saito T, Webster RG, Characterization of a new avian-like influenza A virus from horses in China, Virology 1992, 188(1):245-255.
Horimoto T, Rivera E, Pearson J, Senne D, Krauss S, Kawaoka Y, et al. Origin and molecular changes associated with emergence of a highly pathogenic H5N2 influenza virus in Mexico. Virology 1995; 213:223-30.
Каверин Н.В., Смирнов Ю.А., Межвидовая трансмиссия вирусов гриппа А и проблема пандемий, Вопросы вирусологии, 2003, 3, 4-10.
Toshihiro Ito, Hideo Goto, Eiji Yamamoto, Hiroko Tanaka, Mutsuko Takeuchi, Masaru Kuwayama, Yoshihiro Kawaoka, and Koichi Otsuki, Generation of a Highly Pathogenic Avian Influenza A Virus from an Avirulent Field Isolate by Passaging in Chickens, Journal of Virology, May 2001, Vol. 75, No. 9, p. 4439-4443
Webster RG. Influenza: An Emerging Disease, Emerging Infectious Diseases 1998; 4(3):436-441.
Sang Heui Seo and Robert G. Webster, Cross-Reactive, Cell-Mediated Immunity and Protection of Chickens from Lethal H5N1 Influenza Virus Infection in Hong Kong Poultry Markets, Journal of Virology, Mar. 2001, Vol. 75, No. 6, p. 2516-2525
David E. Swayne, Michael L. Perdue, Joan R. Beck, Maricarmen Garcia1, David L. Suarez, Vaccines protect chickens against H5 highly pathogenic avian influenza in the face of genetic changes in field viruses over multiple years, Veterinary Microbiology 74 (2000) 165-172
D.L. Suarez, S. Schultz-Cherry, Immunology of avian influenza virus: a review, Developmental and Comparative Immunology 24 (2000) 269-283
Avian Flu Virus Linked to Human Conjunctivitis and Fatal ARDS Case, Proc Natl Acad Sci USA 2004; 101:1356-1361
Paul Sockett, Avian influenza, Can Med Assoc J FEB. 10, 1998; 158 (3) 369
Kennedy F. Shortridgea, Peng Gaob, Yi Guanc, Toshihiro Itod, Yoshihiro Kawaokab, Deborah Markwella, Ayato Takadae, Robert G. Webster, Interspecies transmission of influenza viruses: H5N1 virus and a Hong Kong SAR perspective, Veterinary Microbiology 74 (2000) 141-147
Terrence M. Tumpey, David L. Suarez, Laura E. L. Perkins, Dennis A. Senne, Jae-gil Lee, Youn-Jeong Lee, In-Pil Mo, Haan-Woo Sung, and David E. Swayne, Characterization of a Highly Pathogenic H5N1 Avian Influenza A Virus Isolated from Duck Meat, Journal of Virology, June 2002, Vol. 76, No. 12, p. 6344-6355
Masato Hatta and Yoshihiro Kawaoka, The continued pandemic threat posed by avian influenza viruses in Hong Kong, Trends in Microbiology Vol.10 No.7 July 2002, 340-345
Eric C.J. Claas, Pandemic influenza is a zoonosis, as it requires introduction of avian-like gene segments in the human population, Veterinary Microbiology 74 (2000) 133-139
Nopporn Wong-Anan, Human Avian Influenza Cases Confirmed in Thailand
Tan Ee Lyn, Avian Influenza Virus H5N1 May Have Become More Virulent
Emma Hitt, PhD, Avian Flu: What Clinicians Need to Know
David L. Suarez, Evolution of avian influenza viruses, Veterinary Microbiology 74 (2000) 15-27
Lois A. Zitzow, Thomas Rowe, Timothy Morken, Wun-Ju Shieh, Sherif Zaki, and Jacqueline M. Katz, Pathogenesis of Avian Influenza A (H5N1) Viruses in Ferrets, Journal of Virology, May 2002, Vol. 76, No. 9, p. 4420-4429
Jody K. Dybing, Stacey Schultz-Cherry, David E. Swayne, David L. Suarez, and Michael L. Perdue, Distinct Pathogenesis of Hong Kong-Origin H5N1 Viruses in Mice Compared to That of Other Highly Pathogenic H5 Avian Influenza Viruses, Journal of Virology, Feb. 2000, Vol. 74, No. 3, p. 1443-1450
Masato Hatta, Peng Gao, Peter Halfmann, Yoshihiro Kawaoka, Molecular Basis for High Virulence of Hong Kong H5N1 Influenza A Viruses, Science, 7 september 2001, VOL 293, 1840-1842
Peng Gao, Shinji Watanabe, Toshihiro Ito, Hideo Goto, Krisna Wells, Martha McGregor, A. James Cooley, and Yoshihiro Kawaoka, Biological Heterogeneity, Including Systemic Replication in Mice, of H5N1 Influenza A Virus Isolates from Humans in Hong Kong, Journal of Virology, Apr. 1999, Vol. 73, No. 4, p. 3184-3189
Terrence M. Tumpey, Xiuhua Lu, Timothy Morken, Sherif R. Zaki, and Jacqueline M. Katz, Depletion of Lymphocytes and Diminished Cytokine Production in Mice Infected with a Highly Virulent Influenza A (H5N1) Virus Isolated from Humans, Journal of Virology, July 2000, Vol. 74, No. 13, p. 6105-6116
Michael L. Perdue, David L. Suarez, Structural features of the avian influenza virus hemagglutinin that influence virulence, Veterinary Microbiology 74 (2000) 77-86
Mikhail Matrosovich, Nannan Zhou, Yoshihiro Kawaoka, and Robert Webster, The Surface Glycoproteins of H5 Influenza Viruses Isolated from Humans, Chickens, and Wild Aquatic Birds Have Distinguishable Properties, Journal of Virology, Feb. 1999, Vol. 73, No. 2, p. 1146-1155
Erich Hoffmann, Juergen Stech, Irina Leneva, Scott Krauss, Christoph Scholtissek, Po San Chin, Malik Peiris, Kennedy F. Shortridge, and Robert G. Webster, Characterization of the Influenza A Virus Gene Pool in Avian Species in Southern China: Was H6N1 a Derivative or a Precursor of H5N1, Journal of Virology, July 2000, Vol. 74, No. 14, p. 6309-6315
David L. Suarez, Michael L. Perdue, Nancy Cox, Thomas Rowe, Catherine Bender, Jing Huang, and David E. Swayne, Comparisons of Highly Virulent H5N1 Influenza A Viruses Isolated from Humans and Chickens from Hong Kong, Journal of Virology, Aug. 1998, Vol. 72, No. 8, p. 6678-6688
Thomas G. Fanning, Richard D. Slemons, Ann H. Reid, Thomas A. Janczewski, James Dean, and Jeffery K. Taubenberger, 1917 Avian Influenza Virus Sequences Suggest that the 1918 Pandemic Virus Did Not Acquire Its Hemagglutinin Directly from Birds, Journal of Virology, Aug. 2002, Vol. 76, No. 15, p. 7860-7862
Jacqueline M. Katz, Xiuhua Lu, Terrence M. Tumpey, Catherine B. Smith, Michael W. Shaw, and Kanta Subbarao, Molecular Correlates of Influenza A H5N1 Virus Pathogenesis in Mice, Journal of Virology, Nov. 2000, p. 10807-10810 Vol. 74, No. 22
Yuri A. Smirnov, Asya K. Gitelman, Elena A. Govorkova, Aleksandr S. Lipatov, Nikolai V. Kaverin, Influenza H5 virus escape mutants: immune protection and antibody production in mice, Virus Research 99 (2004) 205-208
Rick A. Bright, Ted M. Ross, Kanta Subbarao, Harriet L. Robinson, and Jacqueline M. Katza, Impact of glycosylation on the immunogenicity of a DNA-based influenza H5 HA vaccine, Virology 308 (2003) 270-278
Ayato Takada, Noritaka Kuboki, Katsunori Okazaki, Ai Ninomiya, Hiroko Tanaka, Hiroichi Ozaki, Shigeyuki Itamura, Hidekazu Nishimura, Masayoshi Enami, Masato Tashiro, Kennedy F. Shortridge, and Hiroshi Kida, Avirulent Avian Influenza Virus as a Vaccine Strain against a Potential Human Pandemic, Journal of Virology, Oct. 1999, Vol. 73, No. 10, p. 8303-8307
Xiuhua Lu, Terrence M. Tumpey, Timothy Morken, Sherif R. Zaki, Nancy J. Cox, and Jacqueline M. Katz, A Mouse Model for the Evaluation of Pathogenesis and Immunity to Influenza A (H5N1) Viruses Isolated from Humans, Journal of Virology, July 1999, Vol. 73, No. 7, p. 5903-5911
Janice M. Riberdy, Kirsten J. Flynn, Juergen Stech, Robert G. Webster, John D. Altman, and Peter C. Doherty, Protection against a Lethal Avian Influenza A Virus in a Mammalian System, Journal of Virology, Feb. 1999, Vol. 73, No. 2, p. 1453-1459
Shantha Kodihalli, Hideo Goto, Darwyn L. Kobasa, Scott Krauss, Yoshihiro Kawaoka, and Robert G. Webster, DNA Vaccine Encoding Hemagglutinin Provides Protective Immunity against H5N1 Influenza Virus Infection in Mice, Journal of Virology, Mar. 1999, Vol. 73, No. 3, p. 2094-2098
Terrence M. Tumpey, Mary Renshaw, John D. Clements, and Jacqueline M. Katz, Mucosal Delivery of Inactivated Influenza Vaccine Induces B-Cell-Dependent Heterosubtypic Cross-Protection against Lethal Influenza A H5N1 Virus Infection, Journal of Virology, June 2001, Vol. 75, No. 11, p. 5141-5150
Thomas Rowe, Robert A. Abernathy, Jean Hu-Primmer, William W. Thompson, Xiuhua Lu, Wilina Lim, Keiji Fukuda, Nancy J. Cox, and Jacqueline M. Katz, Detection of Antibody to Avian Influenza A (H5N1) Virus in Human Serum by Using a Combination of Serologic Assays, Journal of Clinical Microbiology, Vol. 37, No. 4, Apr. 1999, p. 937-943
Aleksandr S. Lipatov, Scott Krauss, Yi Guan, Malik Peiris, Jerold E. Rehg, Daniel R. Perez, and Robert G. Webster, Neurovirulence in Mice of H5N1 Influenza Virus Genotypes Isolated from Hong Kong Poultry in 2001, Journal of Virology, Mar. 2003, Vol. 77, No. 6, p. 3816-3823
Taisuke Horimoto and Yoshihiro Kawaoka, Direct Reverse Transcriptase PCR To Determine Virulence Potential of Influenza A Viruses in Birds, Journal of Clinical Microbiology, Mar. 1995, Vol. 33, No. 3, p. 748-751
Lynn A. Cooper and Kanta Subbarao, A Simple Restriction Fragment Length Polymorphism-Based Strategy That Can Distinguish the Internal Genes of Human H1N1, H3N2, and H5N1 Influenza A Viruses, Journal of Clinical Microbiology, July 2000, Vol. 38, No. 7, p. 2579-2583
Erica Spackman, Dennis A. Senne, T. J. Myers, Leslie L. Bulaga, Lindsey P. Garber, Michael L. Perdue, Kenton Lohman, Luke T. Daum, and David L. Suarez, Development of a Real-Time Reverse Transcriptase PCR Assay for Type A Influenza Virus and the Avian H5 and H7 Hemagglutinin Subtypes, Journal of Clinical Microbiology, Sept. 2002, Vol. 40, No. 9, p. 3256-3260
Paul Sockett, Avian influenza, Can Med Assoc J o FEB. 10, 1998; 158 (3) 369
Irina A. Leneva, Olga Goloubeva, Robert J. Fenton, Margaret Tisdale, and Robert G. Webster, Efficacy of Zanamivir against Avian Influenza A Viruses That Possess Genes Encoding H5N1 Internal Proteins and Are Pathogenic in Mammals, Antimicrobial Agents and Chemotherapy, Apr. 2001, Vol. 45, No. 4, p. 1216-1224
Elena A. Govorkova, Irina A. Leneva, Olga G. Goloubeva, Karen Bush, and Robert G. Webster, Comparison of Efficacies of RWJ-270201, Zanamivir, and Oseltamivir against H5N1, H9N2, and Other Avian Influenza Viruses, Antimicrobial Agents and Chemotherapy, Oct. 2001, p. 2723-2732 Vol. 45, No. 10
Influenza Epidemic of 1918 Linked With Avian Virus
G. G. Brownlee and E. Fodor, The predicted antigenicity of the haemagglutinin of the 1918 Spanish influenza pandemic suggests an avian origin, Phil.Trans. R. Soc. Lond. B (2001) 356, 1871-1876
Ryan-Poirier K.A., Kawaoka Y. Distinct glycoprotein inhibitors of influenza A virus in different animal sera, J.Virol., 1991, vol.65, 389-395.
Toshihiro Ito, J. Nelson S. S. Couceiro, Sorge Kelm, Linda G. Baum, Scott Krauss, Maria R. Castrucci, Isabella Donatelli, Hiroshi Kida, James C. Paulson, Robert G. Webster, and Yoshihiro Kawaoka, Molecular Basis for the Generation in Pigs of Influenza A Viruses with Pandemic Potential, Journal of Virology, Sept. 1998, Vol. 72, No. 9, p. 7367-7373
K. S. Li, K. M. Xu, J. S. M. Peiris, L. L. M. Poon, K. Z. Yu, K. Y. Yuen, K. F. Shortridge, R. G. Webster, and Y. Guan, Characterization of H9 Subtype Influenza Viruses from the Ducks of Southern China: a Candidate for the Next Influenza Pandemic in Humans?, Journal of Virology, June 2003, Vol. 77, No. 12, p. 6988-6994
Y. P. Lin, M. Shaw, V. Gregory, K. Cameron, W. Lim, A. Klimov, K. Subbarao, Y. Guan, S. Krauss, K. Shortridgei, R. Webster, N. Cox, and A. Hay, Avian-to-human transmission of H9N2 subtype influenza A viruses: Relationship between H9N2 and H5N1 human isolates, PNAS, August 15, 2000, vol. 97, no. 17, 9654-9658
Angela N. Cauthen, David E. Swayne, Stacey Schultz-Cherry, Michael L. Perdue, and David L. Suarez, Continued Circulation in China of Highly Pathogenic Avian Influenza Viruses Encoding the Hemagglutinin Gene Associated with the 1997 H5N1 Outbreak in Poultry and Humans, Journal of Virology, July 2000, Vol. 74, No. 14, p. 6592-6599
Nan Nan Zhou, Kennedy F. Shortridge, Eric C. J. Claas, Scott L. Krauss, and Robert G. Webster, Rapid Evolution of H5N1 Influenza Viruses in Chickens in Hong Kong, Journal of Virology, Apr. 1999, Vol. 73, No. 4, p. 3366-3374
Marianne L.L. van Genugten, Marie-Louise A. Heijnen, Johannes C. Jager, Pandemic Influenza and Healthcare Demand in the Netherlands: Scenario Analysis, Emerg Infect Dis 9(5), 2003, http://www.medscape.com/viewarticle/453679
Webster RG. Influenza: An Emerging Disease, Emerging Infectious Diseases 1998; 4(3):436-441.
www.cdc.gov
Похожие работы
... . Поэтому пандемии столь опасны и столь смертоносны. 3. Мир может находиться на грани новой пандемии. Она начнется, если произойдет мутация вируса, вызывающего птичий грипп. 4. Ни одна страна мира не может чувствовать себя в безопасности от пандемии. Даже закрытие границ не способно остановить распространение болезни. Ранее она накрывала мир за 6-9 месяцев, ныне, из-за лучшего развития массовых ...
... мед университета имени Д. Галицкого Наталия Виноградова. Она также призвала прекратить рекламу «Арбидола» и «Амиксина». Министерство здравоохранения и социального развития Российской Федерации рекомендует схемы лечения гриппа A/H1N1 для людей с легкими и тяжелыми формами заболевания, а также меры профилактики, сообщает пресс-служба ведомства. Лечение легких случаев гриппа A/H1N1, лечение взрослых ...
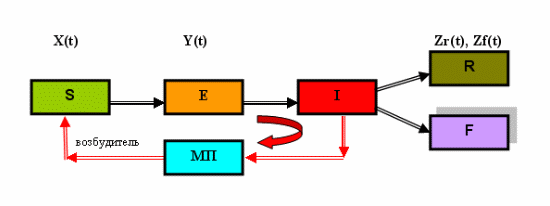
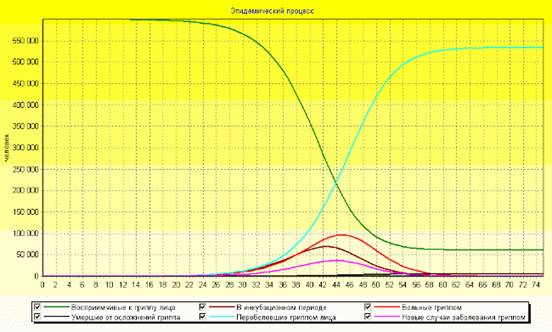
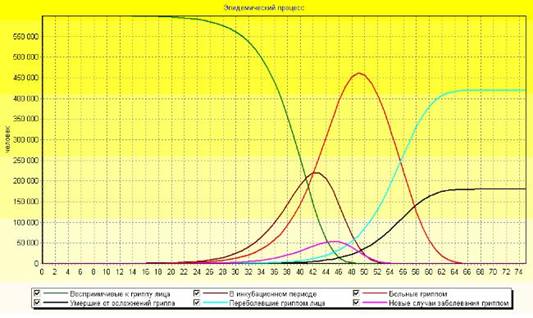
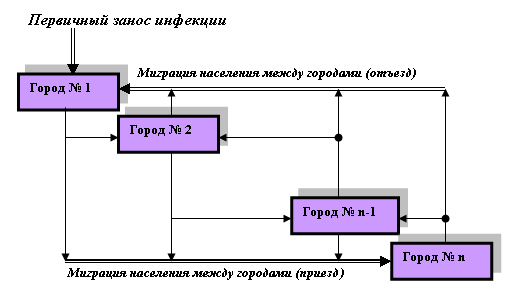
... суперсистемы нелинейных интегро-дифференциальных уравнений 1-7 с соответствующими начальными и граничными условиями, которые записываются для каждого "i"-го города (i=1, 2, 3, …, n). Суперсистема уравнений крупномасштабной эпидемии гриппа в системе из "n" городов страны была реализована в виде компьютерной программы "GRIPP. EXE" (см. рисунок 6), с помощью которой были проведены вычислительные ...
... птиц. Иммунитет, специфическая профилактика После переболевания гриппом птица приобретает нестерильный иммунитет продолжительностью до 6 мес. Наибольшее значение при профилактике высокопатогенного гриппа птиц получили инактивированные вакцины как самые эпизоотологически безопасные. Такие вакцины разработаны и применяются в России. Вакцинированная птица приобретает напряженный иммунитет. ...
0 комментариев