Навигация
Молекулярные биопроцессорные системы для микропрограммной обработки генетической информации
83699
знаков
0
таблиц
1
изображение
8. Молекулярные биопроцессорные системы для микропрограммной обработки генетической информации.
Гены служат только для хранения информации, поэтому её необходимо сначала считывать, а затем определённым образом перерабатывать с тем, чтобы получить форму, приспособленную для непосредственного применения в различных биологических процессах. Вот для этой цели в клетке и применяются аппаратные средства транскрипции и трансляции, которые представляют собой ничто иное, как молекулярные системы для микропрограммной переработки генетической информации. Поэтому фактически каждая живая клетка для микропрограммной обработки генетической информации применяет такие аппаратные устройства, которые с кибернетической точки зрения вполне эквивалентны молекулярным биологическим процессорам.
Известно, что ДНК и РНК живой клетки построены из типовых мономерных звеньев – нуклеотидов. Однако между этими двумя нуклеиновыми кислотами имеются существенные структурные и функциональные различия, которые и привели к появлению в живой клетке особой молекулярной биопроцессорной системы, которая предназначена для считывания информации с ДНК-матрицы и переноса её на структуру РНК. “Этот процесс носит название транскрипции (переписывания). При этом часть двойной спирали ДНК раскручивается, и вдоль одной из её цепей движется особый фермент, который выстраивает нуклеотидные мономеры РНК против их партнёров на цепи ДНК и соединяет эти мономеры друг с другом, так что образуется длинная цепь РНК. На ДНК-матрице образуется три типа РНК: информационная (иРНК), транспортная (тРНК) и рибосомная (рРНК)” [7].
Вспомним, генетическая информация кодируется в виде последовательности нуклеотидов ДНК, – значит, все программы ДНК записаны и хранятся на линейном языке ДНК, а переписываются (транскрибируются) они на “линейную” последовательность нуклеиновых кислот РНК. Известно, что первичный транскрипт РНК в клетках эукариот – это точная копия гена, содержащая как экзоны (кодирующие последовательности транскрипта), так и интроны (некодирующие последовательности), которые должны быть удалены. В блоке процессинга “последовательности интронов вырезаются из середины транскрипта РНК, в результате чего образуется молекула иРНК, непосредственно кодирующая белок. Поскольку кодирующие последовательности с обеих сторон интрона после его удаления соединяются друг с другом, эту реакцию назвали сплайсингом РНК. Сплайсинг РНК протекает в клеточном ядре вдали от рибосом, и РНК переносится в цитоплазму только после завершения этого процесса” [8].
Процесс вырезания интронов и сплайсинг РНК (в операционном блоке процессинга) относится к малоисследованным информационным процессам. Таким образом, задача по считыванию генетической информации в оперативную память структуры РНК решается путем выполнения отдельных микроопераций строго в соответствии с программой того участка ДНК, который определяется структурным геном. А сама генетическая память, транскрипционный аппарат, блок процессинга и каналы ввода/вывода информации представляют собой такое множество сложных молекулярных устройств, совокупность которых действительно представляет собой ядерную биопроцессорную систему управления.
Результатом работы транскрипционной процессорной системы является формирование транспортных и рибосомных РНК, и главное, – загрузка в оперативную память иРНК структурной и программной информации, которая необходима для выполнения различных биологических функций живой клетки. Поэтому основной смысл действия ядерных биопроцессорных систем транскрипции сводится к тому, чтобы передать программную информацию ДНК в оперативную память структуры РНК.
Так происходит считывание генетической информации и передача её к другой биопроцессорной системе для трансляции. В виде информационной РНК, которая в клеточной системе выполняет роль оперативной памяти, генетические программы по синтезу полипептидных цепей передаются к белоксинтезирующему аппарату клетки, то есть к рибосомам. Отметим, что все биопроцессорные системы трансляции генетической информации действуют по одному и тому же принципу, хотя и задействованы в различных управляющих блоках, которые указаны в структурной схеме (блоки катаболизма, биосинтеза элементной базы или биосинтеза различных макромолекул клетки). Разница заключается в различном программном обеспечении иРНК этих блоков.
Далее, в цитоплазме, программная информация переводится (транслируется) с оперативной памяти линейных цепей иРНК на полипептидные цепи белковых молекул. Этот этап характеризуется применением генетического кода и сменой молекулярного носителя информации, когда с помощью молекулярного аппарата трансляции и аминокислотной системы элементов строятся и программируются линейные структуры различных ферментов и других клеточных белков.
Кратко рассмотрим принцип работы молекулярной биопроцессорной системы трансляции (трансляционный аппарат). Основным компонентом типовой биопроцессорной единицы является рибосома. Важная регулирующая роль в синтезе белка помимо иРНК принадлежит тРНК. С помощью специального фермента аминоацил-тРНК-синтетазы тРНК присоединяет на одном из своих концов молекулу соответствующей аминокислоты, в результате чего возникает комплекс аминоацил-тРНК. тРНК при участии белковых факторов устройства управления и энергии гуанозинтрифосфата (ГТФ) доставляет аминокислоту в рибосому для включения ее в растущий полипептид. С помощью своего антикодона тРНК информационно взаимодействует с комплементарным ему кодоном иРНК. Благодаря этой функции тРНК дешифрует генетический код в иРНК-матрице и переводит его в биологический код аминокислотной последовательности белка. Таким образом, обеспечивается необходимая последовательность микроопераций включения аминокислот в синтезируемую полипептидную цепь, строго в соответствии с микропрограммой заданной иРНК. Поэтому функционирование биопроцессорной единицы (рибосомы) в основном состоит из повторяющихся рабочих циклов, каждый из которых соответствует выполнению одной микрокоманды программы иРНК.
Рабочий цикл содержит в общем случае последовательность определенных этапов, которые наглядно отражены в соответствующей биологической литературе. Сама рибосома, в частности, обладает каталитической функцией, ответственной за образование пептидных связей в цепи белка. Как мы видим, иРНК в биопроцессоре играет роль матричного модуля оперативной памяти, несущего микропрограмму преобразования генетической информации в структурную и функциональную информацию полипептидной цепи белка.
Следовательно, задача по преобразованию генетической информации в линейную структуру белка решается путем выполнения отдельных элементарных микроопераций строго в соответствии с заданной микропрограммой, которая заранее была загружена в оперативную память структуры иРНК. При этом системой реализующей процесс трансляции с известными стадиями инициации, элонгации и терминации является молекулярный биологический процессор. Так как молекулярные биопроцессорные единицы (рибосомы) трансляционного аппарата локально рассредоточены по различным блокам и компартментам, то клеточная система управления процессами строится в виде набора единичных процессоров, для которых, как правило, предусматривается своя локальная оперативная память в виде иРНК. Множество подобных молекулярных процессорных единиц обычно соединяются каналами связи, образуя сеть.
Итак, все ядерные биопроцессорные системы различных управляющих систем (катаболизма, биосинтеза элементной базы, биосинтеза макромолекул клетки и т. д., см. структурную схему) обеспечивают транскрипцию (считывание) программной информации с соответствующих участков структуры ДНК и осуществляют загрузку и размещение её в оперативной памяти структур РНК. Далее программная информация оперативной памяти иРНК становится основой работы биопроцессорных систем трансляции, то есть служит для перевода текста программ с языка нуклеиновых кислот на полипептидный язык белковых молекул. При этом осуществляется не только перевод программы с одного языка на другой, но и загрузка программ и данных в структуру белковых молекул. Таким образом, осуществляется перекодирование информации, записанной генетическим кодом и перевод её в программную информацию, записанную кодом аминокислотным.
Заметим, что смена молекулярных носителей обычно сопряжена с процессами кодирования и декодирования информации. К примеру, аминокислотные цепи белков, с одной стороны, являются конечным продуктом процесса декодирования генетической информации, а с другой – они же являются начальным процессом кодирования стереохимической организации белковых молекул с помощью аминокислотного кода. Причем, если генетический код служит для переноса и трансляции генетической программной информации на “линейную” структуру белка, то аминокислотный код является тем молекулярным кодом, с помощью которого осуществляется сначала преобразование, а затем, и, через деятельность белков, – воплощение и реализация генетической информации. Очевидно, что генетическим кодом кодируется только первичная, – “линейная” структура полипептидной цепи. Однако “конкретная конформация (вторичная, третичная и четвертичная структуры) любого белка полностью определяется первичной структурой входящих в его состав полипептидных цепей и зависит от химических свойств боковых групп аминокислотных остатков” [9].
Следовательно, вторичная, третичная и четвертичная структуры белковых молекул кодируются и программируются уже другим молекулярным кодом – аминокислотным. В результате конформационных преобразований и процессинга макромолекула фермента (белка) формирует характерную трехмерную конформацию со своими стереохимическими кодами и, в связи с этим, приобретает свой информационно-кибернетический статус. Весь ход процессинга и адресной доставки фермента в соответствующий операционный блок осуществляется в виде отдельных операций манипуляторами устройства управления, точно в соответствии с кодовыми компонентами белка.
Каждый фермент или другой белок клетки по своим индивидуальным кодам адресации доставляется в свой операционный блок. В операционном блоке, точно в соответствии с функциональным адресным кодом и кодом каталитической операции фермент, действуя как молекулярный биологический автомат, выполняет определенный тип химической реакции. Процесс рецепции информации подлинного субстрата, осуществляемый активным центром фермента, вызывает конформационные изменения в фермент-субстратном комплексе, при которых кодовые химические группы фермента и молекулы субстрата занимают самое оптимальное положение для прохождения каталитической операции.
Важно отметить, что подключение объекта управления (молекулы субстрата), через кодовый стереохимический контакт комплементарного сопряжения, ведёт к индукции электронно-конформационного возбуждения фермент-субстратного комплекса. Присоединение подлинного субстрата сначала ведёт к переброске электронов и протонов между ферментом и молекулой субстрата, усилению электронной перестройки вдоль сопряженной системы связей, что соответственно приводит к возбуждению фермент-субстратного комплекса и, как итог, благодаря подвижным водородным связям, ведёт к динамическим конформационным сдвигам и срабатыванию “силового молекулярного привода” аппарата химического катализа фермента.
Эти механизмы обеспечивают ферменту не только химическую, но и динамическую реактивность и, как результат, – автоматический режим его работы. Возникшие конформационные изменения в фермент-субстратном комплексе сопровождаются разрывом или образованием химических связей субстрата, которые происходят с высвобождением или затратой энергии. В случае необходимости эти процессы поддерживаются химической энергией в форме АТФ. Быстрому протеканию ферментативной реакции способствует высокая химическая и динамическая реактивность фермента.
Высокая химическая реактивность обеспечивается режимом полифункционального катализа, когда на превращаемую химическую связь субстрата одновременно действует стереохимическая комбинация различных каталитически активных химических группировок активного центра (код операции) фермента. Интересным фактом здесь является то, что белковые молекулы стереохимическим способом решают сразу две задачи, – информационной коммуникации и полифункционального катализа.
Динамическая реактивность фермента, при взаимодействии фермента с субстратом, создаёт напряжение, то есть ориентирует и фиксирует взаимодействующие химические группы таким образом, что это создаёт механическую составляющую, которая снижает энергию активации и способствует эффективному прохождению реакции.
Можно считать, что, в рамках сделанных допущений, информационная модель описывает процесс управления химической реакции, ведущий к образованию продуктов реакции. Образование продуктов реакции сопровождается нарушением их физико-химического соответствия управляющим кодовым компонентам фермента, а это приводит к возврату фермента в исходное состояние. Фермент, как взведённая пружина, возвращаясь в исходное состояние, способствует выбросу продуктов реакции из активного центра. Если фермент является аллостерическим, то на него могут воздействовать регуляторные молекулы обратных связей, влияя, таким образом, на ход химической реакции. Так происходит реализация управляющей генетической информации [2].
Заметим также, что клеточная система сразу же получает информацию о ходе управляемых процессов в виде стереохимических кодов продуктов реакции, которые становятся субстратами для других ферментов или выступают в роли молекул обратной связи. Сигнальная (осведомляющая) информация субстратов служит для информирования управляющей системы о состоянии управляемых объектов, о ходе реакций, об эффективности протекающих процессов и т. д. Отличительной особенностью белков клетки является их способность адекватно и сходным образом отвечать на довольно слабые информационные воздействия, достаточно мощными обратимыми конформационными изменениями. В этом, видимо, и заключается основа и сущность их биологической активности.
Способность белка индуцировано возбуждаться и адекватно отвечать на сигнальную информацию изменением своей конформации является специфической особенностью. Конформация фермента меняется при взаимодействии его с субстратом, молекула гемоглобина – при соединении с кислородом, конформационные изменения обеспечивают функционирование сократительных белков и т. д.
Способность ферментов и других белков клетки автоматически отвечать на слабые информационные воздействия, довольно мощными обратимыми конформационными изменениями, используется клеткой практически для всех биологических функций. А этап фермент-субстратного взаимодействия является заключительным фрагментом биокибернетического управления. По всей вероятности, это и есть те, пока недостающие и разыскиваемые фрагменты информационного управления, указывающие на единство процессов управления и информации в каждой живой клетке! Известно также, что некоторые ферменты и белки программно объединяются между собой или с молекулами РНК в агрегатированные автоматы и становятся способными к выполнению сложнейших биологических функций. К молекулярным агрегатам такого рода можно отнести ДНК и РНК-полимеразы, рибосомы, АТФ-синтетазу и т. д.
Здесь мы рассмотрели работу управляющей подсистемы клетки, действие которой непосредственно связано с программной информацией генетической памяти. Очевидно, что нет никаких причин сомневаться в информационной основе рассмотренных выше процессов управления. Теперь нам важно понять сущность управляемых клеточных процессов и убедиться в том, что, несмотря на химическую основу, они также носят информационный характер!
9. Информационная основа управляемых процессов.Одна из отличительных особенностей клеточной системы управления заключается в том, что она информационно взаимодействует с молекулярными объектами управления. Вспомним, – все объекты управления (субстраты), точно так же, как и сама система управления, состоят не только из типовых био-логических элементов (и химических знаков), но и построены по одним и тем же типовым закономерностям. Этот факт позволяет живой клетке не только осуществлять управление превращениями субстратов (или пищевых продуктов), но и осуществлять самоуправление своими же биологическими компонентами.
Очевидно, что все биохимические элементы, а значит и биомолекулы клетки (в том числе и молекулы субстратов), обладают разными типовыми функциональными и боковыми группами, атомами и их химическими связями, которые свободно узнаются и тестируются соответствующими ферментами. Боковые и функциональные атомные группы, атомы и их химические связи – это и есть те опознавательные знаки, благодаря которым управляющая система легко может идентифицировать любой био-логический элемент клетки. Поэтому в живой клетке, кроме молекулярного алфавита различных элементов, существует ещё и свой химический алфавит типовых атомных групп и атомов, манипулируя которыми управляющая система может осуществлять их движение от одного элемента к другому (а, значит, и между молекулами). Поэтому, циркуляция атомных групп и атомов определяет свою субмолекулярную форму движения информации, которая в живой клетке организована в виде управляемых ступенчатых химических реакций! [6].
Таким образом, информационные процессы в живой клетке практически затрагивают не только молекулярный уровень организации, но и, что удивительно, субмолекулярный – атомный! Однако следует отметить, – если целостные элементы в основном служат для организации самих аппаратных устройств и управляющих процессов клетки, то отдельные химические знаки используются не только в качестве информационных сигналов для организации управляемых химических процессов, но применяются и для построения или реорганизации (перекодировки) самих био-логических элементов. В связи с этим, управляющая система клетки, в целом, способна манипулировать различными химическими буквами, символами и знаками, которым предписан определённый биологический и информационный смысл [3].
Очевидно, что все управляемые процессы представляют собой ничто иное, как те ступенчатые химические реакции, которые определяют пути клеточного метаболизма. Только, в ступенчатых химических реакциях различные ферменты способны манипулировать отдельными химическими знаками био-логических элементов. Эта способность управляющей системы основана на том, что при фермент-субстратных взаимодействиях, адресные и операционные коды определенных ферментов соответствуют тем или иным боковым или функциональным атомным группам или атомам и их химическим связям.
Значит, молекулярные коды соответствий, и средства их передачи существуют не только для важнейших систем био-логических элементов – нуклеотидов и аминокислот. Они существуют для любых мономеров и их химических знаков. А одинаковые кодовые комбинации элементов и их боковых атомных групп в активных центрах ферментов всегда воспринимаются конкретной клеткой как одна и та же (эквивалентная) информация, реализуемая в одних и тех же действиях. Очевидно, что для информационного манипулирования различными химическими буквами, символами и знаками живая клетка применяет свои специфические химические или стереохимические молекулярные коды. Специалистам лишь следует научиться их правильно выявлять и идентифицировать.
Кодовые компоненты активных центров ферментов могут специфически (стереохимически и комплементарно) взаимодействовать с доступными для них атомными группами и химическими связями биомолекул (субстратов). Поэтому все субстраты для своих ферментов являются сигнальными молекулами, несущими осведомляющую стереохимическую информацию! На этом основана молекулярная биохимическая логика информационных взаимодействий между ферментами и их субстратами.
В ходе каждой химической реакции, которая управляется своим ферментом, обычно происходит лишь небольшое химическое изменение, например, удаление, перенос или присоединение какого-нибудь атома, боковой или функциональной группы или отдельного биохимического элемента. Иными словами, часть выходного звена управляющего аппарата должна координировать в пространстве и во времени совокупность огромного числа ступенчатых реакций: окисления, восстановления, расщепления, межмолекулярного переноса атомных групп и т. д. Поэтому в качестве объектов управления в клетке могут выступать как отдельные био-логические элементы (нуклеотиды, аминокислоты, простые сахара и жирные кислоты), так и различные биологические молекулы, состоящие из этих элементов, – то есть многочисленные молекулы субстратов.
Каждый объект управления (субстрат) является носителем в “законсервированном” (статическом) виде определённой структурной биологической информации и химической энергии, накопленной в его химических связях. Поэтому все органические питательные вещества, поступающие в живую систему, представляют собой молекулярные информационно-энергетические субстраты, которые поставляют в клетку необходимые структурные, информационные и энергетические компоненты. И всё это клетка получает в результате информационной переработки субстратов (данных). Благодаря стереохимической форме представления информации, сигнальными элементами субстратов для управляющей системы являются лишь те элементы, к которым она в данный момент имеет доступ. Другие же сигнальные элементы (буквы, символы или знаки) временно маскируются в трёхмерной структуре субстрата.
Поэтому информационное преобразование молекулы субстрата, при обработке её различными ферментами, осуществляется последовательно, шаг за шагом (программно), в виде отдельных единичных каталитических операций. Таким образом, все биологические процессы управления и химического превращения веществ в клетке сопряжены с процессами преобразования, как управляющей, так и осведомляющей молекулярной информации. Поскольку каждый фермент способен управлять лишь какую-то одну цепь реакций данного соединения, не влияя на другие возможные реакции, то в отдельно взятом компартменте (операционном блоке) одновременно может протекать множество различных химических реакций.
В связи с этим, можно сделать заключение о том, что других специальных механизмов синхронизирующих работу белков и ферментов, по-видимому, не требуется (кроме сигналов обратных связей или изменения физических и химических факторов микросреды).
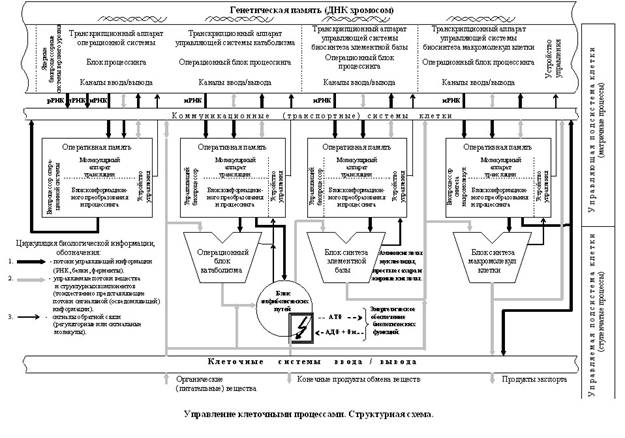
10. Операционные блоки ступенчатых процессов.Ступенчатые биохимические процессы – это деградация или синтез различных простых органических соединений. Это именно тот, программно управляемый биохимический “генератор жизни”, который осуществляет вечное движение органического вещества и энергии и поддерживает баланс разрушительных и созидательных процессов в живой клетке. Очевидно, что управляющая система клетки, по свому назначению, является той информационной системой, которая служит для управления молекулярными биологическими объектами (субстратами). На структурной схеме показаны операционные блоки катаболических и амфиболических (центральных) путей. Важнейшие из них – гликолиз, b -окисление жирных кислот, цикл трикарбоновых кислот и пути распада аминокислот обеспечивают поступление электронов и протонов в электрон-транспортную систему и образование углеродсодержащих соединений (около десяти веществ).
Как видно из структурной схемы, каждый операционный блок содержит свою управляющую и управляемую части. Поступление в блок молекулярных автоматов или манипуляторов – выходного звена управления биопроцессоров, показано жирными черными стрелками. Управляемые потоки вещества, тождественно представляющие потоки сигнальной (осведомляющей) информации субстратов, показаны в виде серых стрелок. Каждый операционный блок предназначен для переработки своей субстратной информации, или, с точки зрения биохимии, для осуществления определенных биохимических реакций. К примеру, блок амфиболических путей обеспечивает не только поступление в блок синтеза элементной базы соответствующих углеродсодержащих соединений, но и осуществляет энергообеспечение живой клетки в форме АТФ. “Благодаря разной локализации ферментов катаболизма и анаболизма эти противоположные метаболические процессы протекают в клетке одновременно. Их связывают центральные, или амфиболические процессы. Примером служит цикл трикарбоновых кислот. Тесная связь между анаболизмом и катаболизмом проявляется на трех уровнях:
1) на уровне источников углерода: продукты катаболизма могут быть исходными субстратами анаболических реакций;
2) на энергетическом уровне: в процессе катаболизма образуются АТФ и другие высокоэнергетические соединения; анаболические процессы протекают с их потреблением; 3) на уровне восстановительных эквивалентов: реакции катаболизма являются в основном окислительными; процессы анаболизма, наоборот, потребляют восстановительные эквиваленты” [10].
Все ступенчатые процессы находятся под генетическим контролем. Сотни протекающих в операционных блоках химических реакций программно организованы с помощью ферментов – молекулярных автоматов, в виде множества различных последовательностей идущих друг за другом операций (реакций). В блоке синтеза молекулярной элементной базы с использованием предшественников идут управляемые процессы биосинтеза аминокислот, нуклеотидов, простых сахаров и жирных кислот.
Из структурной схемы видно, что живая клетка способна сама создавать молекулярную элементную базу для построения своих систем или получать ее из внешней среды. Аминокислоты и нуклеотиды в основном используются в матричных процессах биосинтеза белков и нуклеиновых кислот.
В блоке синтеза структурных и функциональных макромолекул клетки под управлением своих программ идут процессы биосинтеза и процессинга сложных макромолекул белков, ферментов, компонентов мембран и органелл и т. п., которые, после транспортировки, войдут составной частью в клеточные ансамбли, и будут работать в разных клеточных структурах. А создаваемые различными биопроцессорными системами информационные компоненты (рРНК, тРНК, иРНК, белки, ферменты т. д.) – это, по существу, и есть тот программно-аппаратный парк, который работает в различных по своему назначению операционных блоках. Живая клетка на любом отрезке своего развития всегда имеет необходимый и достаточный набор программных и программируемых молекулярно-аппаратных средств, необходимый для обеспечения управления всеми своими химическими процессами и биологическими функциями. Поэтому главной задачей генетической памяти состоит в том, чтобы передать необходимые данные и программную информацию биологическим молекулам и структурам клетки. А загруженная в биомолекулы структурная и программная информация является основой их информационного и функционального поведения в общей системе управления живой клетки.
В связи с этим, все белковые и другие биомолекулы клетки представляют собой не только потоки биоорганического вещества, но они же образуют и информационные потоки и сети, контролирующие различные биохимические и молекулярные функции живой клетки (организма). Программирование этих потоков и сетей обеспечивается экспрессией десятков и сотен различных генов, объединённых между собой скоординированными управляющими и регуляторными воздействиями. А если учесть, что различные ферментативные системы, состоящие порой из десятков и сотен ферментов, участвуют в организации множества различных последовательностей идущих друг за другом химических реакций, которые в совокупности составляют клеточный метаболизм, то можно констатировать, что управление химическими процессами и биологическими функциями клетки осуществляется молекулярными информационными потоками и сетями “автоматизированного” управления.
Отсюда, как следствие, вытекает и тот факт, что все клеточные процессы управляются, регулируются и взаимно координируются той программной информацией, которая (с помощью генетической информации и элементной базы) загружена в аппаратную систему клетки, то есть находится в функциональных биомолекулах и структурах клетки! Каждая из управляющих систем клетки (операционная система, управляющие системы катаболизма, биосинтеза элементной базы, биосинтеза макромолекул клетки и т. д.), состоящая из генетической памяти, комплекса локальных биопроцессорных устройств и выходного управляющего звена – молекулярных биологических автоматов и манипуляторов, воспринимает информацию о ходе химических превращений, об эффективности протекающих процессов, об изменении внешних и внутренних физических и химических факторов и, в зависимости от результата, корректирует управляющие воздействия.
Системы запрограммированы на четкое взаимодействие управляющих и управляемых подсистем (и друг с другом), на оптимальное прохождение всех биохимических и энергетических процессов. Живая клетка в любое время имеет необходимый и достаточный набор программных, энергетических и функциональных средств для поддержания и обновления своих структур, обеспечения процессов развития и жизнедеятельности.
В настоящее время накоплен огромнейший экспериментальный и теоретический материал по изучению живой материи, который рассредоточен по различным областям и дисциплинам молекулярных биологических наук.
Теперь уже стало очевидным, что чрезвычайная детализация изучаемых биохимических процессов ведёт только к бесконечной лавине все новых и новых проблем и вопросов. Это не приводит ни к осознанию сущности живой материи, ни к пониманию причин и механизмов её функционирования. По мнению автора статьи, в молекулярной биологии наступила пора переосмысления имеющихся знаний и сведений в пользу новой, альтернативной науки – молекулярной биологической информатики. О необходимости такого шага свидетельствует вся история возникновении и развития жизни на Земле, которая повсеместно связана с наследственной информацией и действующей на её основе универсальной мультипроцессорной молекулярно-биологической системы управления живой клетки.
1. А. Ленинджер. Основы биохимии. Пер. с англ. в 3-х томах – М: Мир, 1985.
2. Ю. Я. Калашников. Ферменты и белки живой клетки – это молекулярные биологические автоматы с программным управлением. Дата публикации: 30 июня 2006г., источник: SciTecLibrary.ru; Сайт: , дата публикации: 13.12.2006г.
3. Ю. Я. Калашников. Кодирование и программирование биологических молекул. Дата публикации: 01.01.2007г., источник: http://new-idea.kulichki.com/
4. Ю. Я. Калашников. Единство вещества, энергии и информации – основной принцип существования живой материи. Дата публикации: 30 июня 2006г., источник: SciTecLibrary.ru; Сайт: http://new-idea.kulichki.com/, дата публикации: 07.12.2006г.
5. Ю. Я. Калашников. Аспекты молекулярной биохимической логики и информатики. Дата публикации: 05.12.2006г., источник: http://new-idea.kulichki.com/
6. Ю. Я. Калашников. Концепция информационной молекулярно-биологической системы управления. – М., 2005.–88с. – Депонир. в ВИНИТИ РАН 14.04.05, №505-В2005
7. П. Кемп, К. Армс. Введению в биологию. Пер. с англ. – М: “Мир”, 1988.
8. Б. Альбертс, Д. Брей и другие. Молекулярная биология клетки. Пер. с англ., Том 2 – М: “Мир”, 1994.
9. Ф. Айала, Дж. Кайгер. Современная генетика. Пер. с англ. в 3-х томах – М: Мир, 1988.
10. А. А. Анисимов, А. Н. Леонтьева и др. Основы биохимии. – “Высшая школа”, 1986. Дата публикации: 5 марта 2
Похожие работы
... появившиеся информационные макромолекулы, предвестники жизни, стали на длительный путь формирования биологических структур. Можно без преувеличения сказать, что химический способ представления информации стал именно тем гениальным изобретением природы, с помощью которого была подведена черта под химической эволюцией материи, и были открыты необъятные дали и непредсказуемые пути великой эволюции – ...
... зарождения. Это, по всей вероятности, и есть та диктатура информационной субстанции, которая определяет нашу биологическую сущность на самом фундаментальном – молекулярном уровне. 4. Информация правит нашим миром или информационная концепция эволюции. Информация, – она кажется нам нереальной и неопределимой. Необъятный мир её разнообразен и не изучен. Но она не только существует, но даже живёт ...
... автоматически отвечать на слабые информационные воздействия, довольно мощными обратимыми конформационными изменениями, используется клеткой практически для всех биологических функций. 4. Ферменты и белки – это молекулярные биологические автоматы с программным управлением. В живой клетке имеется множество локально рассредоточенных объектов управления (субстратов). Для эффективного управления ими ...
... за две-три недели. Во время обследований необходимо избегать нарушений в работе других видов транспорта четкой координацией управления ими. Изучение пассажиропотоков позволяет выявить основные закономерности их колебания для использования результатов обследований в планировании и организации перевозок. Иначе говоря, характер изменения пассажиропотоков на маршрутах и в целом по конкретному ...
0 комментариев