Навигация
Ингибирование ферментативной активности
25178
знаков
0
таблиц
4
изображения
Министерство сельского хозяйства РФ ФГОУ ВПО "Оренбургский государственный аграрный университет" КУРСОВАЯ РАБОТА по биохимии на тему: Ингибирование ферментативной активности


Оренбург – 2010
Содержание
1. Ингибиторы ферментов. Типы ингибирования активности ферментов
1.1 Обратимое ингибирование
1.1.1 Конкурентное ингибирование
1.1.2 Неконкурентное ингибирование
1.1.3 Бесконкурентное ингибирование
1.2 Необратимое ингибирование
1.3 Аллостерическое ингибирование
2. Новый вид ингибирования ферментативной активности
3. Применение ингибиторов ферментов
ЗАКЛЮЧЕНИЕ
Список использованной литературы1. Ингибиторы ферментов. Типы ингибирования активности ферментов
Известно, что активность ферментов сравнительно легко может быть уменьшена с помощью разнообразных воздействий. Такое снижение скорости ферментативных реакций принято называть торможением активности, или ингибированием ферментов.
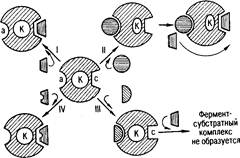
Рис 1. Схема активирования и ингибирования действия фермента (по Ю. Б. Филипповичу): а. – аллостерический центр фермента; К - каталитический центр; с - субстратный центр
Ферменты являются белками, соответственно их активность можно снизить или полностью ликвидировать путем воздействий, приводящих к денатурации белков (нагревание, действие концентрированных кислот, щелочей, солей тяжелых металлов и т.п.) Это неспецифическое подавление активности ферментов, имеющее значение при изучении ферментативных реакций, не представляет особого интереса для исследования их механизма. Гораздо большее значение имеет исследование ингибирования с помощью веществ, специфически и обычно в небольших количествах взаимодействующих с ферментами – ингибиторов ферментов. Расшифровка механизмов многих биологических процессов, таких как гликолиз, цикл Кребса и других, стала возможной лишь в результате применения специфических ингибиторов различных ферментов (Н.Е. Кучеренко, Ю.Д. Бабенюк и др., 1988).
Некоторые ингибиторы ферментов являются для организма животных и человека эффективными лекарственными веществами, другие — смертельными ядами (В.П. Комов, В.Н. Шведова, 2004).
Ингибиторы взаимодействуют с активными центрами молекулы фермента, инактивируя функциональные группы белков. Они могут взаимодействовать с металлами, входящими в состав молекул ферментов и фермент-субстратных комплексов, инактивируя их. Высокие концентрации ингибиторов разрушают четвертичную, третичную и вторичную структуры молекулы фермента, вызывая его денатурацию (А.И. Кононский, 1992).
Недавно открыты антиферменты (антиэнзимы, или антизимы), представляющие собой белки, действующие как ингибиторы ферментов. К подобным веществам относятся, например, ингибитор трипсина, обнаруженный в соевых бобах, и сывороточный антитрипсин. Недавно открыт в печени животных антифермент орнитиндекарбоксилазы. Антизимы, вероятнее всего, образуют труднодиссоциируемые комплексы с соответствующими ферментами, выключая их из химических реакций. Иногда ингибитор является составным компонентом предшественника фермента, или входит в состав сложных комплексов ферментов. Однако до сих пор не выяснено, являются ли подобные антиферменты истинными ингибиторами или регуляторными субъединицами.
Если ингибитор вызывает стойкие изменения пространственной третичной структуры молекулы фермента или модификацию функциональных групп фермента, то такой тип ингибирования называется необратимым. Чаще, однако, имеет место обратимое ингибирование, поддающееся количественному изучению на основе уравнения Михаэлиса-Ментен. Обратимое ингибирование в свою очередь разделяют на конкурентное и неконкурентное
На практике многие ингибиторы не проявляют тех свойств, которые характерны для чисто конкурентного или чисто неконкурентного ингибирования. Другой способ классификации ингибиторов основывается на характере места их связывания. Одни из них связываются с ферментом в том же месте, что и субстрат (в каталитическом центре), а другие — на значительном расстоянии от активного центра (в аллостерическом центре) (Р. Марри, Д. Греннер и др., 1993).
1.1 Обратимое ингибирование Различают три типа обратимого ингибирования ферментов: конкурентное, неконкурентное и бесконкурентное, в зависимости от того, удается или не удается преодолеть торможение ферментативной реакции путем увеличения концентрации субстрата. 1.1.1 Конкурентное ингибированиеКонкурентный ингибитор конкурирует с субстратом за связывание с активным центром, но в отличие от субстрата связанный с ферментом конкурентный ингибитор не подвергается ферментативному превращению. Отличительная особенность конкурентного ингибирования состоит в том, что его можно устранить или ослабить, просто повысив концентрацию субстрата. Например, если при заданных концентрациях субстрата и конкурентного ингибитора активность фермента подавлена на 50 %, то мы можем уменьшить степень ингибирования, повысив концентрацию субстрата.
По своей трехмерной структуре конкурентные ингибиторы обычно напоминают субстрат данного фермента. Благодаря такому сходству конкурентному ингибитору удается "обмануть" фермент и связаться с ним. Конкурентное ингибирование можно количественно изучать на основе теории Михаэлиса-Ментен. Конкурентный ингибитор I просто обратимо присоединяется к ферменту Е, образуя с ним комплекс
Е + I = EI.
Конкурентное ингибирование проще всего можно распознать экспериментальным путем, определив влияние концентрации ингибитора на зависимость начальной скорости реакции от концентрации субстрата. Для выяснения вопроса о том, по какому типу - конкурентному или неконкурентному - происходит обратимое ингибирование фермента, используют метод двойных обратных величин. Из графиков, построенных в двойных обратных координатах, можно определить также значение константы диссоциации комплекса фермент ингибитор (см. рис. 1) (А. Ленинджер, 1985)
Конкурентное ингибирование может быть вызвано веществами, имеющими структуру, похожую на структуру субстрата, но несколько отличающуюся от структуры истинного субстрата. Такое ингибирование основано на связывании ингибитора с субстратсвязывающим (активным) центром (см. рис. 2).
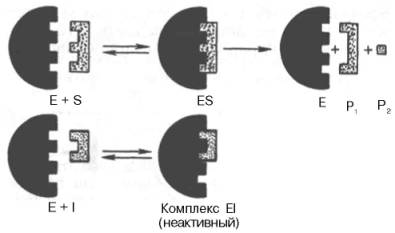
Рис. 2. Общий принцип конкурентного ингибирования (схема по В.Л. Кретовичу). Е - фермент; S - субстрат; Р1 и Р2 - продукты реакции; I - ингибитор.
В качестве примера можно привести действие малоновой кислоты на реакцию, которая катализируется сукцинатдегидрогеназой и связана с превращением янтарной кислоты в фумаровую. Добавление малоновой кислоты к реакционной смеси снижает или полностью останавливает ферментативную реакцию, так как она является конкурентным ингибитором сукцинатдегидрогеназы. Сходства малоновой кислоты с янтарной достаточно для образования комплекса с ферментом, однако распад этого комплекса не происходит. При увеличении концентрации янтарной кислоты она вытесняет малоновую кислоту из комплекса, в результате активность сукцинатдегидрогеназы восстанавливается.
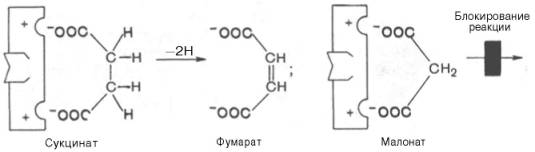
Рис. 3. Конкурентное ингибирование реакции превращения янтарной кислоты в фумаровую под действием малоновой кислоты.
Структуры субстрата (сукцинат) и ингибитора (малонат) все же несколько различаются. Поэтому они конкурируют за связывание с активным центром, и степень торможения будет определяться соотношением концентраций малоната и сукцината, а не абсолютной концентрацией ингибитора. Таким образом, ингибитор может обратимо связываться с ферментом, образуя фермент-ингибиторный комплекс. Этот тип ингибирования иногда называют ингибированием по типу метаболического антагонизма (см. рис. 3).
В общей форме реакция взаимодействия ингибитора с ферментом может быть представлена следующим уравнением:
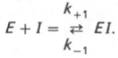
Образовавшийся комплекс, называемый фермент-ингибиторным комплексом ЕI, в отличие от фермент-субстратного комплекса ES не распадается с образованием продуктов реакции.
Многие лекарственные вещества ингибируют ферменты человека и животных по конкурентному типу. Например, что для лечения некоторых инфекционных заболеваний, вызываемых бактериями, применяют сульфаниламидные препараты. Оказалось, что эти препараты имеют структурное сходство с парааминобензойной кислотой, которую бактериальная клетка использует для синтеза фолиевой кислоты, являющейся составной частью ферментов бактерий. Благодаря этому структурному сходству сульфаниламид блокирует действие фермента путем вытеснения парааминобензойной кислоты из комплекса с ферментом, синтезирующим фолиевую кислоту, что ведет к торможению роста бактерий.
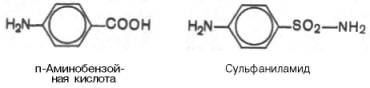
В структуру пептидогликана клеточной стенки бактерий включен D-аланин, отсутствующий в организме животных и человека. Для синтеза клеточной стенки бактерии при помощи фермента аланин-рацемазы превращают L-аланин животных в D-форму. Аланин-рацемаза характерна для бактерий и не обнаружена у млекопитающих. Следовательно, она представляет хорошую мишень для ингибирования лекарственными препаратами. Замещение одного из протонов метильной группы на фтор дает фтораланин, с которым связывается аланин-рацемаза, что приводит к ее ингибированию.
Некоторые аналоги витамина В6 и фолиевой кислоты, в частности дезоксипиридоксин и аминоптерин, действуют как конкурентные, так называемые коферментные, ингибиторы (или антивитамины), тормозящие многие интенсивно протекающие при патологии биологические процессы в организме. Применение подобных аналогов в медицинской практике (в частности, в дерматологии и онкологии) основано на конкурентном вытеснении коферментов из субстратсвязывающих центров ключевых ферментов обмена (Т.Т Березов, Б.Ф. Коровкин, 1990).
Таким образом, можно конструировать лекарственные вещества, ингибирующие ферменты по конкурентному типу. Чтобы быть эффективным, ингибитор должен иметь высокое сродство к ферменту. В противном случае необходимо назначать большие дозы лекарственных препаратов, чтобы активно конкурировать с эндогенным субстратом за активный центр фермента (В.П. Комов, В.Н. Шведова, 2004).
1.1.2 Неконкурентное ингибированиеНеконкурентное ингибирование тоже обратимо, но не может быть ослаблено или устранено повышением концентрации субстрата
В случае неконкурентного ингибирования ингибитор присоединяется к ферменту не в активном центре, где связывается субстрат, а совсем в другом месте. При этом конформация молекулы фермента изменяется таким образом, что происходит обратимая инактивация его каталитического центра. Неконкурентные ингибиторы связываются обратимо как со свободным ферментом, так и с комплексом ES, образуя неактивные комплексы EI и ESI.
Неконкурентное ингибирование вызывается веществами, не имеющими структурного сходства с субстратами и часто связывающимися не с активным центром, а в другом месте молекулы фермента. Степень торможения во многих случаях определяется продолжительностью действия ингибитора на фермент. При данном типе ингибирования благодаря образованию стабильной ковалентной связи фермент часто подвергается полной инактивации, и тогда торможение становится необратимым (Н.З. Хазипов, А.Н. Аскарова, 2001).
Использование графиков, построенных в двойных обратных координатах для анализа данных, полученных при исследовании ингибирования ферментативных реакций, позволяет легко отличать конкурентные ингибиторы от неконкурентных.
Наиболее важные неконкурентные ингибиторы представляют собой образующиеся в живых организмах промежуточные продукты метаболизма, способные обратимо связываться со специфическими участками на поверхности некоторых регуляторных ферментов и изменять при этом активность их каталитических центров. Примером может служить ингибирование L-треониндегидратазы L-изолейцином (А. Ленинджер, 1985).
Примером необратимого ингибирования является действие йодацетата, ДФФ, а также диэтил-n-нитрофенилфосфата и солей синильной кислоты. Это действие заключается в связывании и выключении функциональных групп или ионов металлов и молекуле фермента.
Следует указать, что неконкурентное ингибирование также может быть обратимым и необратимым, поскольку отсутствует конкуренция между субстратом и ингибитором за активный центр. Примеры необратимого ингибирования приведены ранее. При обратимом неконкурентном ингибировании субстрат S и ингибитор I связываются с разными центрами, поэтому появляется возможность образования как комплекса EI, так и тройного комплекса EIS; последний может распадаться с освобождением продукта, но с меньшей скоростью, чем комплекс ES.
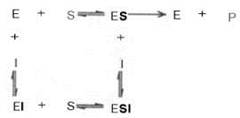
Этот тип неконкурентного ингибирования чаще всего наблюдается у ферментов, катализирующих превращения более одного субстрата, когда связывание ингибитора не блокирует связывание субстрата с активным центром. Ингибитор при этом соединяется как со свободным ферментом, так и с ES-комплексом (Т.Т. Березов, Б.Ф. Коровкин, 1990).
1.1.3 Бесконкурентное ингибированиеВ редких случаях степень торможения активности фермента может увеличиваться с повышением концентрации субстрата. Для этого типа торможения был предложен термин "бесконкурентное ингибирование".
Бесконкурентное ингибирование имеет место, когда ингибитор взаимодействует с ферментом только в составе фермент-субстратного комплекса, препятствуя его распаду (Уэбб Л., 1966).
Примером необратимого действия ингибиторов на ферменты могут служить фосфорорганические вещества, применяемые в качестве инсектицидов.
Известно, кроме того, так называемое бесконкурентное ингибирование, когда ингибитор связывается с ферментом также в некаталитическом центре, однако не со свободным ферментом, а только с ES-комплексом в виде тройного комплекса ESI (Т.Т. Березов, Б.Ф. Коровкин, 1990).
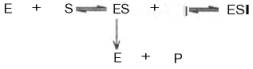
При бесконкурентном ингибировании ингибитор связывается только с фермент-субстратным комплексом, но не со свободным ферментом. Субстрат, связываясь с ферментом, изменяет его конформацию, что делает возможным связывание с ингибитором. Ингибитор, в свою очередь, так меняет конформацию фермента, что катализ становится невозможным
Один из механизмов такого торможения обусловлен возможностью соединения ингибитора с комплексом ES с образованием неактивного или медленно реагирующего тройного комплекса EIS (Т.Т. Березов, Б.Ф. Коровкин, 1990).
Ингибирование субстратом — частный случай бесконкурентного ингибирования, когда две молекулы субстрата связываются с ферментом, что препятствует образованию продукта
Активность многих ферментов тормозится избытком субстрата, причем имеется несколько механизмов этого процесса.
1) Если в образовании фермент-субстратного комплекса участвует несколько функциональных групп фермента, то возможно одновременное присоединение к активному центру двух или более субстратов, что однозначно приведет к образованию неактивного комплекса.
2) В случае избытка субстрата возможно его присоединение не только к активному центру, но и к другим химическим группировкам, функционально связанным с активным центром. Такого рода взаимодействие может помешать ферментативной реакции.
3) Увеличение концентрации субстрата может повысить ионную силу реакционной среды и, как следствие, затормозить скорость ферментативной реакции.
Торможение продуктами реакции связано с тем, что они могут связываться с ферментом или с каким-либо другим компонентом системы таким образом, что скорость прямой реакции снижается.1.2 Необратимое ингибирование Необратимые ингибиторы связывают или разрушают функциональную группу молекулы фермента, необходимую для проявления его каталитической активности. Происходит формирование стабильного комплекса ингибитора с ферментом, ведущее к его необратимой инактивации. Случай необратимого ингибирования можно обнаружить по тому признаку, что при разбавлении раствора не происходит повышения удельной активности фермента, как в случае обратимого ингибирования.
Примером необратимого ингибитора может служить соединение диизопропилфторфосфат (ДФФ), которое ингибирует фермент ацетилхолинэстеразу, играющий важную роль в передаче нервных импульсов. Ацетилхолинэстераза катализирует гидролиз ацетилхолина, функционирующего в качестве нейромедиатора в определенных отделах нервной системы. Ацетилхолин выделяется стимулированной нервной клеткой в синапс, т.е. место соединения одного нейрона с другим. В синапсе ацетилхолин связывается с рецепторами следующего нейрона, вынуждая его проводить нервный импульс. Однако прежде чем второй импульс будет передан через синапс следующему нейрону, ацетилхолин, выделившийся после первого импульса, должен быть гидролизован ацетилхолинэстеразой в месте соединения нервных клеток. Продукты его распада - ацетат и холин - не способны действовать как нейромедиаторы. Необратимый ингибитор ДФФ, обладающий высокой реакционной способностью, присоединяется к гидроксильной группе остатка серина в активном центре ацетилхолинэстеразы, что приводит к образованию каталитически неактивного производного. В результате фермент перестает функционировать. ДФФ, одно из первых отравляющих веществ нервнопаралитического действия, в опытах на животных вызывает нарушение некоторых функций вследствие того, что пораженные нейроны утрачивают способность проводить нервные импульсы. Однако ДФФ обладает и полезными свойствами. На его основе был создан ряд относительно нетоксичных для людей и животных инсектицидов, например малатион. Сам по себе малатион неактивен и в организме высших животных разлагается на продукты, которые считаются безвредными. В организме же насекомых малатион превращается под действием ферментов в активный ингибитор их собственной ацетилхолинэстеразы.
Выяснилось, что ДФФ ингибирует целый класс ферментов, многие из которых способны катализировать гидролиз пептидов или эфирных связей. К этим ферментам относится не только ацетилхолинэстераза, но и трипсин, химотрипсин, эластаза, фосфоглюкомутаза и коконаза (фермент, выделяемый личинкой тутового шелкопряда и используемый ею для гидролиза шелковых нитей и освобождения из кокона). Характерная особенность всех ферментов, ингибируемых ДФФ, состоит в том, что они содержат в активном центре остаток серина, принимающий участие в каталитическом акте
Другой необратимый ингибитор некоторых ферментов, иодацетамид может взаимодействовать с сульфгидрильными (—SH) группами остатков цистеина или с имидазольными группами остатков гистидина, содержащихся в активных центрах этих ферментов. С помощью таких ингибиторов было установлено, что гидроксильная группа серина, тиоловая группа цистеина и имидазольная группа гистидина участвуют в каталитической активности ферментов, принадлежащих к разным классам.
1.3 Аллостерическое ингибированиеАллостерические ингибиторы связываются с отдельными участками фермента вне активного центра. Такое связывание влечет за собой конформационные изменения в молекуле фермента, которые приводят к уменьшению его активности. Аллостерические эффекты встречаются практически только в случае олигомерных ферментов. Кинетику таких систем нельзя описать с помощью простой модели Михаэлиса-Ментен.
Так, йодацетат IСН2—СООН, его амид и этиловый эфир, пара-хлормеркурибензоат ClHg—С6Н4—СООН и другие реагенты сравнительно легко вступают в химическую связь с некоторыми SH-группами ферментов. Если такие группы имеют существенное значение для акта катализа, то добавление подобных ингибиторов приводит к полной потере активности фермента:
R-SH + IСН2—СООН —> НI + R—S—CH2—COOH
Действие ряда других ферментов (холинэстераза, трипсин и химотрипсин) сильно тормозится некоторыми фосфорорганическими соединениями, например ДФФ, вследствие блокирования ключевой гидроксильной группы серина в активном центре (Северин Е.С., 2004).
Похожие работы
... образовательным стандартом Высшего профессионального образования для студентов, обучающихся по специальности 020208 – «Биохимия». 4. Место дисциплины в профессиональной подготовке студентов Курс «кинетики и термодинамики ферментативных реакций» является заключительным курсом в цикле спецдисциплин федерального компонента, посвященных изучению ферментов. Он имеет основополагающее значение, ...
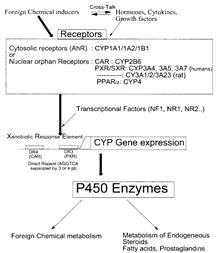
... гем- содержащих монооксигеназ цитохрома P 450, с которыми связываются, подверженные окислительному метаболизму ксенобиотики. [Clin. Pharmcokinet 38, 493, 2000]. Таблица 6 - Механизмы индукции различных изоформ цитохрома Р-450 Механизм индукции Индукция и регуляция P450 Транскрипця генов посредством рецепторов 1A1 (цитозоль AhR), 1A2, 1B1 2A6, 2B6 (CAR), 2C8, 2C9, 2C18, 2C19 3A4, 3A5 ...
... среду [Менщикова, 2008]. 3.2. Анализ содержание GSH и активность глутатионзависимых ферментов в эритроцитах крови практически здоровых людей, проживающих в различных по уровню загрязнения районах г.Красноярска Город Красноярск административно разделен на 7 районов, различаемых по уровню техногенной нагрузки, которая определяется структурой промышленности и энергетики, исходного и ...
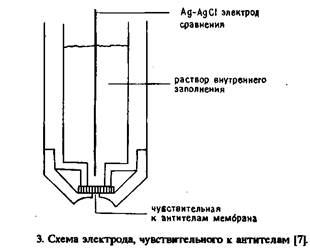
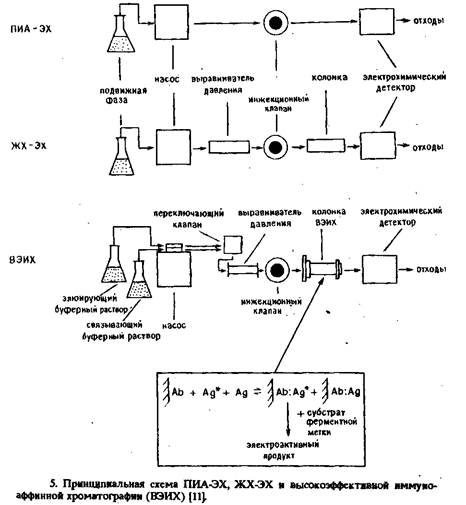
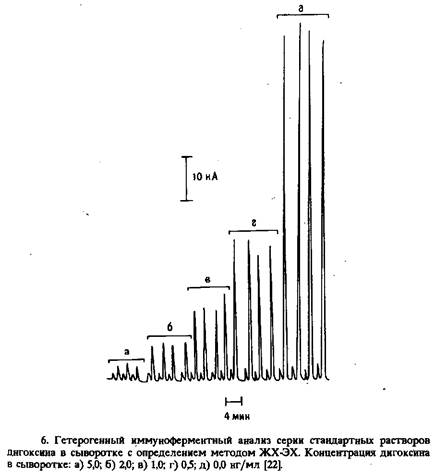
... электрода, обусловленное адсорбцией белков, и перекрестные реакции антител практически не влияют на результат анализа. Неферментативные электроактивные метки. Многообещающим направлением в амперометрическом иммуноанализе представляется применение металлсодержащих соединений. К таким меткам предъявляются следующие требования: 1) хорошие электрохимические свойства, 2) нечувствительность к рН в ...
0 комментариев